
Isolation and characterization of pterosin derivatives from P. aquilinum
Deriváty pterosinu byly izolovány z celých rostlin P. aquilinum. Pro zkoumání fytochemických složek z kapradiny pterosinu byl vodný extrakt postupně rozdělen ethylacetátem (EtOAc) a n-butanolem (n-BuOH) (obr. 1). Opakovaná sloupcová chromatografie frakce rozpustné v EtOAc vedla k izolaci devíti derivátů, včetně (2R)-pterosinu B (1), pterosinu Z (2), (2S)-pterosinu P (3), (3R)-pterosin D (4), (2S)-pterosin A (5), (2S,3R)-pterosin C (6), (2R,3R)-pterosin C (7), (2R)-pterosid B (8) a pterosid Z (9), s čistotou vyšší než 97 % (obr. 1). 2). Opakovaná kolonová chromatografická separace frakce rozpustné v n-BuOH vedla k izolaci šesti derivátů, včetně (2S)-pterosidu A (10), (2S)-pterosidu A2 (11), (2S,3R)-pterosidu C (12), (2R,3R)-pterosidu C (13), (3S)-pterosidu D (14) a (2S)-pterosidu P (15), s čistotou vyšší než 97 %. Struktury sloučenin byly identifikovány na základě analýzy spektrálních dat, včetně MS, 1D- a 2D-NMR (Doplňková informace 1).

Struktury sloučenin pterosinu 1-15xxx
Inhibiční aktivita derivátů pterosinu vůči BACE1, AChE, a BChE
Pro vyhodnocení antiAD potenciálu byla inhibiční aktivita jednotlivých pterosinových sloučenin vůči BACE1 a cholinesterasám hodnocena pomocí příslušných in vitro inhibičních testů (tabulka 1). Všechny testované pterosinové deriváty vykazovaly na koncentraci závislou inhibiční aktivitu vůči BACE1 s rozmezím hodnot IC50 (polo-maximální inhibiční koncentrace) 9,74-94,4 μM, s výjimkou (2S)-pterosinu A a (2S)-pterosidu P, které byly v testovaných koncentracích neaktivní, ve srovnání s IC50 kvercetinu použitého jako pozitivní kontrola, která byla 18,8 μM. Inhibiční účinnost nejsilnějších inhibitorů byla v pořadí (2R,3R)-pterosid C, (3S)-pterosid D, (2R)-pterosid B, (2S,3R)-pterosin C, (2R,3R)-pterosin C, (2S,3R)-pterosid C a (2R)-pterosin B s hodnotami IC50 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 a 29,6 μM. Následně jsme testovali inhibiční potenciál derivátů pterosinu vůči AChE. Všechny testované sloučeniny vykazovaly významnou inhibiční aktivitu vůči AChE s hodnotami IC50 v rozmezí 2,55-110 μM ve srovnání s IC50 vůči AChE berberinu použitého jako pozitivní kontrola, která byla 0,39 μM. Sloučeniny pterosinu, které vykazovaly nejsilnější inhibiční aktivitu vůči AChE, byly (2R)-pterosid B, (2R,3R)-pterosid C, (2S,3R)-pterosid C, (2S,3R)-pterosin C a (2R)-pterosin B s hodnotami IC50 2,55, 3,77, 9,17, 12,8 a 16,2 μM. Nakonec jsme testovali inhibiční schopnosti derivátů pterosinu vůči BChE. Všechny testované pterosinové sloučeniny vykazovaly inhibiční aktivitu vůči BChE s hodnotami IC50, které se pohybovaly od 5,29 do 119 μM, s výjimkou (3R)-pterosinu D, který byl v testovaných koncentracích neaktivní ve srovnání s IC50 berberinu vůči BChE, která byla 3,32 μM. Sloučeniny pterosinu, které vykazovaly nejsilnější inhibiční aktivitu vůči BChE, byly (2R,3R)-pterosid C a pterosid Z s hodnotami IC50 5,29 a 5,31 μM.
Všechny testované pterosinové deriváty vykazovaly významnou inhibiční aktivitu proti BACE1, AChE a BChE současně. Přítomnost další 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolové skupiny jako u pterosidových derivátů významně zvyšovala inhibiční aktivity vůči enzymům. Navíc přítomnost další hydroxymethylové skupiny v poloze 2 indanonového kruhu (2R)-pterosinu B jako u (2S)-pterosinu A nebo methylové skupiny jako u (3R)-pterosinu D a pterosinu Z snižovala inhibiční aktivity vůči enzymům. Zejména přítomnost hydroxymethylové skupiny v poloze 5 indanonového kruhu jako u (2S)-pterosinu P snižovala inhibiční aktivitu vůči BACE1.
Kinetické parametry inhibice enzymů deriváty pterosinu
Ve snaze vysvětlit způsob enzymatické inhibice derivátů pterosinu jsme provedli kinetickou analýzu BACE1 a cholinesteras pro reprezentativní inhibitory (tabulka 2, doplňková informace 2). Nízké Ki (inhibiční konstanta) ukazuje na těsnější vazbu na enzym a účinnější inhibitor. Celkově hodnoty Ki sloučenin korelovaly s příslušnými hodnotami IC50. Inhibice BACE1 sloučeninami (2R,3R)-pterosid C, (3S)-pterosid D a (2R,3R)-pterosin C byla smíšeného typu s hodnotami Ki 12,6, 16,5 a 27. V případě sloučenin (2R,3R)-pterosid D a (2R,3R)-pterosin C byla inhibice BACE1 smíšeného typu.6 µM, zatímco inhibice (2R)-pterosidem B, (2S,3R)-pterosinem C a (2R)-pterosinem B byla nekompetitivní s hodnotami Ki 23,1, 33,8 a 38,3 µM. Inhibice AChE (2R)-pterosidem B, (2R,3R)-pterosidem C, (2R)-pterosinem B, (2S,3R)-pterosinem C a (3S)-pterosidem D byla smíšeného typu s hodnotami Ki 4.89, 8,13, 12,1, 16,3 a 23,1 µM, zatímco (2R,3R)-pterosid C byl inhibitorem nekompetitivního typu s hodnotou Ki 29,6 µM. Inhibice BChE (2R,3R)-pterosinem C, (2R,3R)-pterosidem C, (3S)-pterosidem D a (2R)-pterosidem B byla smíšeného typu s hodnotami Ki 4,77, 9,62, 19.7 a 22,6 µM, zatímco (2S,3R)-pterosin C a (2R)-pterosin B byly nekompetitivní inhibitory s hodnotami Ki 29,9 a 53,5 µM. Tyto výsledky tedy naznačují, že specifické deriváty pterosinu mohou být účinnými inhibitory BACE1, AChE a BChE.
Simulace molekulárního dokování pro BACE1, AChE a BChE
Pro BACE1 a cholinesterázy je k dispozici několik krystalových struktur. Vybrali jsme lidské PDB na základě struktur divokého typu, kokrystalizovaných ligandů a rozlišení struktur. Pro dokování byly vybrány rentgenové krystalové struktury BACE1 komplexované s QUD (kód PDB: 2WJO, rozlišení: 2,5 Å)33, AChE komplexované s E2020 (kód PDB: 4EY7, rozlišení: 2,35 Å)34 a BChE komplexované s 3F9 (kód PDB: 4TPK, rozlišení: 2,70 Å)35 . Zpočátku byly QUD, E2020 a 3F9 extrahovány z krystalových struktur a dokovány do aktivních míst BACE1, AChE a BChE. Následně byly dokovány (2R,3R)-pterosid C, (3S)-pterosid D, (2R,3R)-pterosid C, (2R)-pterosid B, (2S,3R)-pterosid C a (2R)-pterosid B se známým mechanismem inhibice vůči BACE1, AChE a BChE, aby se určila jejich SAR. Výsledky dokování jsou shrnuty v tabulce 3. SAR vybraných inhibitorů smíšeného typu a nekompetitivních inhibitorů BACE1, AChE a BChE umožnilo vyhodnotit nové inhibice enzymů pterosiny zaměřené na vazebné místo.
Náš způsob dokování E2020 byl v souladu s experimentálně stanoveným způsobem vazby, který byl dříve uveden u rekombinantní lidské AChE (rhAChE) (Doplňková informace 3)34. Střední kvadratická odchylka (RMSD) mezi krystalovou a dokovanou konformací E2020 byla 0,54 Å, což naznačuje spolehlivost našeho dokovacího nastavení při reprodukci experimentálního vazebného režimu. Dokovaný mód E2020 navíc vedl k podobné interakci jako u rhAChE-E2020. V naší studii byly během dokování z krystalové struktury odstraněny molekuly vody, proto v této studii nebyly analyzovány interakce zprostředkované vodou. Stejně tak dokované módy QUD a 3F9 byly v souladu s dostupnými experimentálními údaji pro BACE1 33, respektive BChE35 (doplňková informace 3). RMSD mezi krystalovou a dokovanou konformací QUD a 3F9 byly 0,46, resp. 0,60 Å. Dále byla vazebná místa inhibitorů pterosinu ve shodě s předchozí dokovací studií, která zahrnovala BACE1, AChE a BChE38. V této studii však byla použita Tetronarce californica AChE (kód PDB: 1ACJ), která obsahuje mírně odlišné počty zbytků než lidská AChE v důsledku rozdílů v jejich sekvencích.
Dokování BACE1
Na základě typu inhibice a aktivity byly vybrány (2R,3R)-pterosid C a (2R)-pterosid B, aby demonstrovaly dokovací režimy inhibitorů BACE1 smíšeného typu a nekompetitivních inhibitorů. Na obrázcích 3a, b jsou zobrazeny dokovací modely (2R,3R)-pterosidu C a (2R)-pterosidu B. Interakce dokovaných sloučenin uvnitř aktivního místa BACE1 jsou zobrazeny na obr. 4.

Molekulární dokovací modely pro a inhibitor BACE1 smíšeného typu (2R,3R)-pterosid C (modrá barva) a b nekompetitivní inhibitor BACE1 (2R)-pterosid B (hnědá barva). Dokované polohy jsou superponovány na rentgenovou krystalovou strukturu QUD (žlutá barva) (kód PDB: 2WJO). BACE1, zbytky aktivního místa a sloučeniny jsou znázorněny stužkovými, čárovými a tyčkovými modely. Barvy tečkovaných čar vysvětlují typy různých interakcí: interakce vodíkové vazby (zelená), hydrofobní interakce (růžová) a π-sigma interakce (fialová). BACE1 β-site amyloid precursor protein cleaving enzyme 1

Diagram interakcí ligandů a QUD, b (2R,3R)-pterosid C, c (3S)-pterosid D, d (2R,3R)-pterosin C, e (2R)-pterosid B, f (2S,3R)-pterosin C a g (2R)-pterosin B v aktivním místě BACE1. Barvy tečkovaných čar vysvětlují typy různých interakcí: interakce vodíkové vazby (zelená), hydrofobní interakce (růžová) a π-sigma interakce (fialová). BACE1 β-site amyloidového prekurzorového proteinu štěpícího enzymu 1
Dokovaná pozice QUD vykazovala vazebnou energii (B.E) -7,59 kcal/mol. Jak ukazuje obr. 4a, NH2 skupina na chinazolinovém kruhu ligandu vykazovala dvě vodíkové vazby s CO skupinami Asp32 a Asp228 ve vzdálenostech 1,86, resp. 2,17 Å. Třetí vodíková vazba byla pozorována mezi druhou NH skupinou ligandu a CO skupinou Gly230 ve vzdálenosti 2,16 Å. Leu30, Tyr71, Phe108 a Val332 zprostředkovávaly hydrofobní interakce. Na obrázcích 4b-d jsou zobrazeny dokované polohy (2R,3R)-pterosidu C, (3S)-pterosidu D a (2R,3R)-pterosinu C (inhibitory BACE1 smíšeného typu). Byly umístěny ve vazebné kapse lemované Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 a Thr231. Podle úrovně aktivity vykazovaly (2R,3R)-pterosid C (IC50 = 9,74 µM), (3S)-pterosid D (IC50 = 10,7 µM) a (2R,3R)-pterosin C (IC50 = 26,2 µM) B.E. -7,27, -6,93 a -4,84 kcal/mol. (2R,3R)-pterosid C vykazoval vyšší účinnost než (2R,3R)-pterosin C díky existenci další 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolové skupiny, která vytvořila tři vodíkové vazby (obr. 4b). OH skupina hydroxymethylu a 3-OH skupina tetrahydro-pyran-triolového kruhu vykazovaly dvě vodíkové vazby s CO skupinou Asp228 ve vzdálenostech 2,63, resp. 2,12 Å. Kromě toho vykazovala 3-OH skupina vodíkovou vazbu s Thr231 ve vzdálenosti 1,89 Å. V případě (3S)-pterosidu D (obr. 4c) přítomnost 2,2-dimethylové skupiny na indanonovém kruhu mírně změnila vazebné interakce ve srovnání s (2R,3R)-pterosidem C. 3-OH skupina tetrahydro-pyran-triolového kruhu vykazovala pouze jednu vodíkovou vazbu s Asp228 ve vzdálenosti 1,93 Å. Skupina 3-OH indanonového kruhu však vykazovala další vodíkovou vazbu s Ile126 ve vzdálenosti 2,34 Å. Tyto interakce mírně snižovaly aktivitu (3S)-pterosidu D ve srovnání s (2R,3R)-pterosidem C. Dokovací interakce (2R,3R)-pterosidu C zobrazené na obr. 4d ukazují, že 3-OH skupina indanonového kruhu je vázána na Ser36 a Asn37 prostřednictvím dvou vodíkových vazeb ve vzdálenostech 1,83 a 2,07 Å, v tomto pořadí. Ala39, Val69, Trp76, Ile118 a Arg128 byly zapojeny do hydrofobních interakcí, zatímco Val69 vykazoval π-sigma interakci.
Dokované pozice (2R)-pterosidu B, (2S,3R)-pterosinu C a (2R)-pterosinu B (nekompetitivních inhibitorů BACE1) jsou zobrazeny na obr. 4e-g, v tomto pořadí. Byly zakotveny do dutiny uzavřené Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 a Ile126. V souladu s hodnotami jejich aktivity vykazovaly (2R)-pterosid B (IC50 = 18,0 µM), (2S,3R)-pterosin C (IC50 = 23,1 µM) a (2R)-pterosin B (IC50 = 29,6 µM) B.E. -6,16, -5,07 a -4,64 kcal/mol. Jak je zobrazeno na obr. 4e, (2R)-pterosid B vykazoval vyšší aktivitu než (2S,3R)-pterosin C a (2R)-pterosin B díky přítomnosti další 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolové skupiny, která vykazovala čtyři interakce vodíkové vazby. Dvě vodíkové vazby byly pozorovány mezi 4-OH skupinou tetrahydro-pyran-triolového kruhu a NH a CO skupinou Asn37 ve vzdálenostech 2,48 a 2,19 Å. Dále 3- a 5-OH skupiny vykazovaly další dvě vodíkové vazby s Ile126 a Trp76 ve vzdálenostech 2,19 a 2,46 Å. (2S,3R)-pterosin C vykazoval o něco lepší aktivitu než (2R)-pterosin B díky přítomnosti další OH skupiny v poloze 3 indanonového kruhu, která vytvořila vodíkovou vazbu s Lys107 ve vzdálenosti 2,12 Å (obr. 4f). Ostatní interakce byly podobné jako u (2R)-pterosinu B. Jak ukazuje obr. 4g, (2R)-pterosin B vykazoval hydrofobní interakce s Val69, Tyr76 a Phe108.
Dokování AChE
(2R)-pterosid B a (2R,3R)-pterosin C byly vzhledem ke své aktivitě a typu inhibice AChE vybrány jako zástupci pro demonstraci dokovacích režimů inhibitorů smíšeného, respektive nekompetitivního typu. Obrázky 5a, b znázorňují dokovací modely (2R)-pterosidu B a (2R,3R)-pterosinu C v uvedeném pořadí. Interakce dokovaných sloučenin uvnitř aktivního místa AChE jsou zobrazeny na obr. 6.

Molekulární dokovací modely pro a inhibitor AChE smíšeného typu (2R)-pterosid B (modrá barva) a b nekompetitivní inhibitor AChE (2R,3R)-pterosin C (hnědá barva). Dokované polohy jsou superponovány na rentgenovou krystalovou strukturu E2020 (žlutá barva) (kód PDB: 4EY7). AChE, zbytky aktivního místa a sloučeniny jsou znázorněny stužkovými, čárovými a tyčkovými modely. Barvy tečkovaných čar vysvětlují typy různých interakcí: interakce vodíkové vazby (zelená) a hydrofobní interakce (růžová). AChE acetylcholinesterasa

Diagram ligandových interakcí a E2020, b (2R)-pterosidu B, c (2R,3R)-pterosidu C, d (2S,3R)-pterosinu C, e (2R)-pterosinu B, f (3S)-pterosidu D a g (2R,3R)-pterosinu C v aktivním místě AChE. Barvy tečkovaných čar vysvětlují typy různých interakcí: interakce vodíkové vazby (zelená), hydrofobní interakce (růžová) a π-sigma interakce (fialová). AChE acetylcholinesterasa
Dokovaná pozice E2020 prokázala B.E. -10,28 kcal/mol. Jak je znázorněno na obr. 6a, CO skupina indanonového kruhu vytvořila vodíkovou vazbu s NH skupinou Phe295 ve vzdálenosti 1,70 Å. Trp286 a Tyr341 byly zapojeny do π-sigma interakcí, zatímco Trp86, Trp286, Tyr337, Phe338 a Tyr341 zprostředkovávaly hydrofobní interakce. Obrázky 6b-f ukazují dokované polohy (2R)-pterosidu B, (2R,3R)-pterosidu C, (2S,3R)-pterosinu C, (2R)-pterosinu B a (3S)-pterosidu D (inhibitory AChE smíšeného typu), v tomto pořadí. Byly umístěny v aktivním místě obklopeném Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 a Tyr341. V souladu s hodnotami jejich aktivity jsou (2R)-pterosid B (IC50 = 2,55 µM), (2R,3R)-pterosid C (IC50 = 3,77 µM), (2S,3R)-pterosin C (IC50 = 12.8 µM), (2R)-pterosin B (IC50 = 16,2 µM) a (3S)-pterosid D (IC50 = 27,4 µM) vykazovaly B.E. -7,90, -7,49, -6,03, -5,76 a -4,91 kcal/mol. (2R)-pterosid B vykazoval vyšší účinnost než (2S,3R)-pterosin C a (2R)-pterosin B díky přítomnosti další 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolové skupiny, která vytvořila tři interakce vodíkové vazby (obr. 6b). OH skupina hydroxymethylu a 3-OH skupina tetrahydro-pyran-triolového kruhu vykazovaly vodíkové vazby s CO skupinou Asn87 ve vzdálenostech 2,23, resp. 2,28 Å. Dále 4-OH skupina vytvořila vodíkovou vazbu s CO skupinou Trp86 ve vzdálenosti 2,13 Å. V případě (2R,3R)-pterosidu C (obr. 6c) methylová skupina v poloze 2 indanonového kruhu nevykazovala hydrofobní interakci s Tyr341, a proto vykazovala relativně nižší aktivitu než (2R)-pterosid B. Tyr341 si však zachoval hydrofobní interakci s druhou částí indanonového kruhu, jak se ukázalo u (2R)-pterosidu B. Vyšší aktivita (2S,3R)-pterosinu C než (2R)-pterosinu B byla přisuzována existenci další OH skupiny v poloze-3 indanonového kruhu, která vytvořila vodíkovou vazbu s Phe295 ve vzdálenosti 1,82 Å (obr. 6d). Ostatní interakce byly srovnatelné s interakcemi (2R)-pterosinu B. Jak ukazuje obr. 6e, OH skupina hydroxyethylové skupiny v poloze 6 indanonového kruhu vytvořila vodíkovou vazbu s Tyr124 ve vzdálenosti 2,48 Å. Na hydrofobních interakcích se podílely Tyr124, Trp286, Tyr337, Phe338 a Tyr341. Ve srovnání s (2R)-pterosidem B a (2R,3R)-pterosidem C vykazoval (3S)-pterosid D odlišné vazebné interakce v důsledku přítomnosti 2,2-dimethylové skupiny na indanonovém kruhu (obr. 6f). 2,2-dimethylová skupina významně přispěla k odlišné dokovací pozici (3S)-pterosidu D. 5-OH skupina tetrahydro-pyran-triolového kruhu vytvořila vodíkovou vazbu s Asp74 ve vzdálenosti 1,96 Å. CO skupina indanonového kruhu vykazovala vodíkovou vazbu s Tyr72 ve vzdálenosti 2,97 Å. Tyto interakce byly příčinou nízké aktivity (3S)-pterosidu D.
Dokovaná póza (2R,3R)-pterosinu C (nekompetitivní inhibitor AChE) je zobrazena na obr. 6g. (2R,3R)-pterosin C (IC50 = 23,2 µM) vykazoval B.E. -5,01 kcal/mol. Vazebná kapsa (2R,3R)-pterosinu C zahrnovala Tyr72, Trp286, Ser293, Phe297 a Tyr341 se dvěma interakcemi s vodíkovou vazbou. Jedna z vodíkových vazeb byla vytvořena mezi 3-OH skupinou indanonového kruhu a CO skupinou Ser293 ve vzdálenosti 2,03 Å. Druhá vodíková vazba byla pozorována mezi OH skupinou hydroxyethylové skupiny přítomné v poloze 6 indanonového kruhu a Tyr72 ve vzdálenosti 1,94 Å. Rezidua jako Tyr72, Trp286, Phe297 a Tyr341 se účastnila hydrofobních interakcí.
Dokování BChE
S ohledem na úroveň aktivity a typ inhibice BChE byly vybrány (2R,3R)-pterosid C a (2S,3R)-pterosin C, aby demonstrovaly dokovací způsoby smíšeného, resp. nekompetitivního typu inhibitorů BChE. Obrázek 7a, b znázorňuje dokovací modely (2R,3R)-pterosidu C a (2S,3R)-pterosinu C v uvedeném pořadí. Interakce dokovaných sloučenin uvnitř aktivního místa BChE jsou uvedeny na obr. 8.
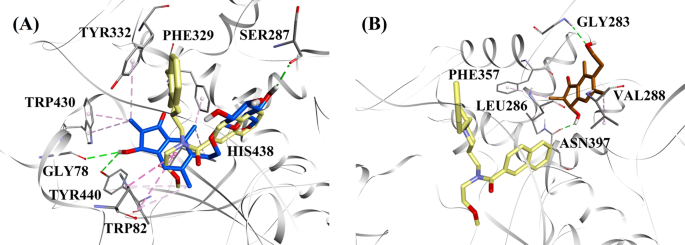
Molekulární dokovací modely pro a inhibitor BChE smíšeného typu (2R,3R)-pterosid C (modrá barva) a b nekompetitivní inhibitor BChE (2S,3R)-pterosin C (hnědá barva). Dokované polohy jsou superponovány na rentgenovou krystalovou strukturu 3F9 (žlutá barva) (kód PDB: 4TPK). BChE, zbytky aktivního místa a sloučeniny jsou znázorněny stužkovými, čárovými a tyčkovými modely. Barvy tečkovaných čar vysvětlují typy různých interakcí: interakce vodíkové vazby (zelená) a hydrofobní interakce (růžová). BChE butyrylcholinesterasa
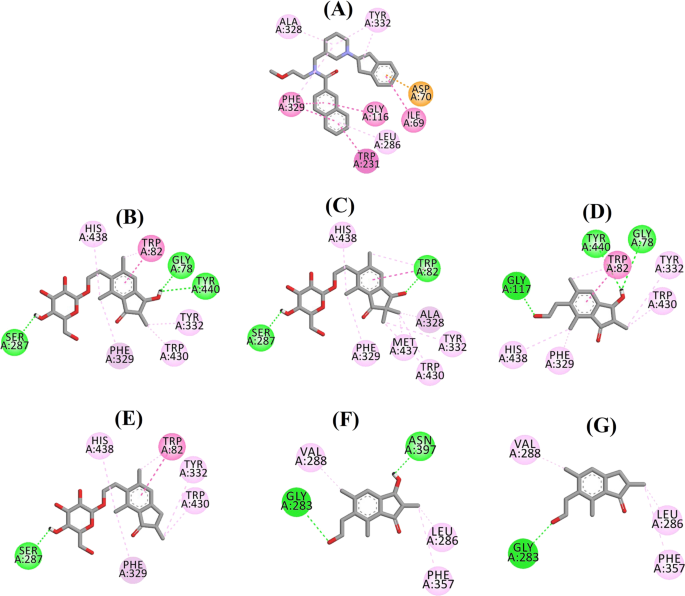
Diagram ligandových interakcí a 3F9, b (2R,3R)-pterosidu C, c (3S)-pterosidu D, d (2R,3R)-pterosinu C, e (2R)-pterosidu B, f (2S,3R)-pterosinu C a g (2R)-pterosinu B v aktivním místě BChE. Barvy tečkovaných čar vysvětlují typy různých interakcí: interakce vodíkové vazby (zelená), hydrofobní interakce (růžová) a π-aniontové interakce (zlatá). BChE butyrylcholinesterasa
Dokovaná póza 3F9 vykazovala B.E. -8,49 kcal/mol. Jak je zobrazeno na obr. 8a, za vazbu ligandu byly zodpovědné především hydrofobní interakce. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 a Tyr332 se podílely na hydrofobních interakcích, zatímco Asp70 vykazoval π-aniontovou interakci. Obrázky 8b-e znázorňují dokované polohy (2R,3R)-pterosidu C, (3S)-pterosidu D, (2R,3R)-pterosinu C a (2R)-pterosidu B (inhibitory BChE smíšeného typu), v tomto pořadí. Jejich vazebnou kapsu tvořily Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 a Tyr440. V souladu s úrovní své aktivity vykazovaly (2R,3R)-pterosid C (IC50 = 5,29 µM), (3S)-pterosid D (IC50 = 19,3 µM), (2R,3R)-pterosin C (IC50 = 20,3 µM) a (2R)-pterosid B (IC50 = 62,0 µM) B.E. -7,23, -6,59, -6,52 a -4,38 kcal/mol. Jak ukazuje obr. 8b, 3-OH skupina indanonového kruhu (2R,3R)-pterosidu C vykazovala dvě vodíkové vazby s Gly78 a Tyr440 ve vzdálenostech 2,87 a 2,89 Å, v tomto pořadí. V případě (3S)-pterosidu D ovlivňovala vazebné interakce 3OH skupiny 2,2-dimethylová skupina na indanonovém kruhu (obr. 8c). Skupina 3-OH vytvořila pouze jednu vodíkovou vazbu s Trp82 ve vzdálenosti 2,94 Å, což mělo za následek relativně nižší aktivitu (3S)-pterosidu D než (2R,3R)-pterosidu C. Jak ukazuje obr. 8d, (2R,3R)-pterosid C nevytvořil vodíkovou vazbu se Ser287 kvůli absenci 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolové skupiny. V důsledku toho poskytoval nižší aktivitu než (2R,3R)-pterosid C. Bylo zjištěno, že absence OH skupiny v poloze 3 indanonového kruhu je zodpovědná za velmi nízkou aktivitu (2R)-pterosidu B (obr. 1). 8e), který nevytvářel vodíkové vazby s Gly78 a Tyr440 podobně jako (2R,3R)-pterosid C.
Dokované polohy (2S,3R)-pterosinu C a (2R)-pterosinu B (nekompetitivních inhibitorů BChE) jsou zobrazeny na obr. 8f, resp. g. Tyto dokované pozice byly obsaženy v dutině uzavřené Gly283, Leu286, Val288, Phe357 a Asn397. Podle úrovně jejich aktivity vykazovaly (2S,3R)-pterosin C (IC50 = 44,3 µM) a (2R)-pterosin B (IC50 = 48,1 µM) B.E. -5,40 a -5,06 kcal/mol. (2S,3 R)-pterosin C byl účinnější než (2R)-pterosin B díky přítomnosti další OH skupiny v poloze 3 indanonového kruhu, která vytvořila vodíkovou vazbu s CO skupinou Asn397 ve vzdálenosti 2,05 Å (obr. 8f). Další interakce byly zjištěny podobné jako u (2R)-pterosinu B. Jak ukazuje obr. 8g, OH skupina v hydroxyethylové skupině v poloze-6 indanonového kruhu vytvořila vodíkovou vazbu s Gly283 ve vzdálenosti 2,40 Å. Zbytky Leu286, Val288 a Phe357 se účastnily hydrofobních interakcí.
Inhibitory smíšeného typu se vážou jak na volný enzym, tak na komplex enzym-substrát, což naznačuje, že se tyto sloučeniny mohou vázat na katalytické místo každého příslušného enzymu. Nekompetitivní inhibitory se vážou na alosterické místo volného enzymu nebo komplexu enzym-substrát. Nedávná studie naznačila, že kompetitivní, smíšené a nekompetitivní inhibitory obsazují různá místa ve vazebných kapsách BACE1, AChE a BChE38. Při dokování pro hodnocení inhibičního mechanismu derivátů pterosinu byla vazebná místa sloučenin definována podle jejich typu inhibice. Výsledky dokování ukázaly, že vazebná místa inhibitorů smíšeného a nekompetitivního typu pro BACE1, AChE a BChE se částečně překrývají v každém odpovídajícím aktivním místě a jsou v souladu s předchozí zprávou38.
Propustnost BBB
PAMPA-BBB, in vitro test umělé membránové propustnosti pro BBB, je jedním z nejspolehlivějších nástrojů fyzikálně-chemického screeningu v rané fázi objevování léčiv zaměřených na CNS40. Systém PAMPA-BBB modeluje transcelulární pasivní difúzi chemických látek přes BBB a měří striktně pasivní transportní mechanismy přes umělou lipidovou membránu na efektivní permeabilitu (Pe, cm/s). Na základě vzorce stanoveného pro predikci permeace BBB byly sloučeniny klasifikovány na (i) „CNS+“ (vysoká predikovaná permeace BBB); Pe (10-6 cm/s) > 4,00, (ii) „CNS-“ (nízká predikovaná permeace BBB); Pe (10-6 cm/s) < 2,00 a (iii) „CNS+/-“ (nejistá permeace BBB); Pe (10-6 cm/s) od 4,00 do 2,00. V souladu s tím vykazovaly (2R)-pterosin B, (2S)-pterosin P a (2S)-pterosin A vysokou permeaci BBB s hodnotami Pe 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s a 6,26 × 10-6 cm/s (tabulka 4, doplňková informace 4). Hodnota Pe (2R)-pterosinu B byla 1,7krát vyšší než hodnota Pe CNS léčiva verapamilu (Pe = 34,6 × 10-6 cm/s), který byl použit jako pozitivní kontrola. (2S,3R)-pterosin C a (2R,3R)-pterosin C vykazovaly nejistý průnik BBB s hodnotami Pe 2,34, resp. 1,98. (2R,3R)-pterosid C, (3S)-pterosid D a (2R)-pterosid B, které mezi testovanými deriváty pterosinu vykazovaly nejsilnější inhibiční aktivitu vůči BACE1 a cholinesteráze, vykazovaly velmi nízkou prostupnost BBB. Existence 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolové skupiny jako u pterosidových derivátů vedla k pozoruhodnému snížení propustnosti BBB. Ve srovnání s (2R)-pterosinem B další přítomnost OH skupiny v poloze 3 indanonového kruhu jako u pterosinu C, hydroxymethylové skupiny v poloze 2 indanonového kruhu jako u (2S)-pterosinu A nebo hydroxymethylové skupiny v poloze 5 indanonového kruhu jako u (2S)-pterosinu P rovněž významně snížila propustnost BBB. Vzhledem k mimořádně vysoké propustnosti BBB a významné inhibici BACE1, AChE a BChE může mít (2R)-pterosin B potenciál vykazovat silnou antiAD aktivitu.
Vliv (2R)-pterosinu B a (2R,3R)-pterosidu C na sekreci peptidů Aβ neuronálními buňkami
Zkoumání funkce (2R)-pterosinu B a (2R,3R)-pterosidu C při snižování vylučování Aβ z neuronálních buněk jsme použili myší neuroblastomovou buněčnou linii, která stabilně nadměrně exprimuje lidský APPswe. Tato buněčná linie představuje buněčný model AD charakterizovaný nadměrným vylučováním Aβ40 a Aβ42. Toxické amyloidní oligomery se tvoří ze dvou izoforem peptidu Aβ s různou délkou. Aβ40 je nejhojnější izoforma Aβ v mozku41 , zatímco Aβ42 se u některých forem AD výrazně zvyšuje42. Sandwichová ELISA Aβ40 ukázala, že (2R)-pterosin B významně snižuje množství peptidu Aβ40 vylučovaného z neuroblastomových buněk do média až o 50 % při 500 μM (P < 0,01) (obr. 9a). Podobně se významně snížila sekrece peptidu Aβ42 neuroblastomovými buňkami v přítomnosti 500 μM (2R,3R)-pterosidu C (P < 0,05) (obr. 9b). Závěrem lze říci, že (2R)-pterosin B a (2R,3R)-pterosid C významně snížily sekreci peptidů Aβ z neuroblastomových buněk při koncentraci 500 μM.

a Účinek (2R)-pterosinu B na sekreci peptidů Aβ. Neuroblastomové buňky vylučující APPswe byly ošetřeny 12, 60, 250 nebo 500 µM (2R)-pterosinu B po dobu 24 h a kondicionovaná média byla odebrána v přítomnosti inhibitoru proteázy. Jako pozitivní kontrola byl použit β-SI (10 µM). Negativní kontrola zahrnovala buňky kultivované v nepřítomnosti testovaných sloučenin. Kvantitativní analýza vylučovaných Aβ40 a Aβ42 v kondicionovaných médiích byla provedena pomocí sendvičové ELISA. Jsou uvedeny průměry ± SD ze tří nezávislých experimentů. Vylučované peptidy Aβ40 se významně snížily v přítomnosti 500 µM (2R)-pterosinu B. CON negativní kontrola, β-SI inhibitor β-sekretázy III, Aβ β-amyloid. *P < 0,05, **P < 0,01. b Vliv (2R,3R)-pterosidu C na sekreci peptidů Aβ. Neuroblastomové buňky vylučující APPswe byly ošetřeny 12, 60, 125, 250 nebo 500 µM (2R,3R)-pterosidu C po dobu 24 h a kondicionovaná média byla odebrána v přítomnosti inhibitoru proteázy. Jako pozitivní kontrola byl použit β-SI (10 µM). Negativní kontrola zahrnovala buňky kultivované v nepřítomnosti testovaných sloučenin. Kvantitativní analýza vylučovaných Aβ40 a Aβ42 v kondicionovaných médiích byla provedena pomocí sendvičové ELISA. Jsou uvedeny průměry ± SD ze tří nezávislých experimentů. Vylučovaný peptid Aβ42 se významně snížil v přítomnosti 500 µM (2R,3R)-pterosidu C. CON negativní kontrola, β-SI inhibitor β-sekretázy III, Aβ β-amyloid. *P < 0.05
Citotoxicita pterosinových derivátů na základě MTT testu43
Obecně pterosinové deriváty vykazovaly zanedbatelnou cytotoxicitu vůči různým normálním a nádorovým buněčným liniím, jako je lidský neuroblastom SH-SY5Y, potkaní gliové buňky C6, myší embryofibroblasty NIH3T3 a myší melanom B16F10, s hodnotami LD50 nad 0 %.5 mM (doplňková informace 5). Zejména (2R)-pterosin B, (3R)-pterosin D, (2S)-pterosin P, (2S)-pterosid A, (2R)-pterosid B a (2R,3R)-pterosid C nevykazovaly cytotoxicitu vůči testovaným buněčným liniím s hodnotami LD50 nad 5 mM. Zajímavé je, že několik pterosinů vykazovalo relativní antiproliferační účinky proti neuronálním buňkám SH-SY5Y ve srovnání s gliovými buňkami C6 mezenchymálního původu. Současné výsledky byly v souladu s předchozí zprávou, která naznačovala, že deriváty pterosinu jsou pro člověka netoxické26.
.