

Příspěvek solného můstku k celkové stabilitě ke složenému stavu proteinu lze posoudit pomocí termodynamických údajů získaných ze studií mutageneze a technik nukleární magnetické rezonance. Pomocí mutovaného proteinu pseudo-wildova typu specificky mutovaného tak, aby zabraňoval vysrážení při vysokém pH, lze určit příspěvek solného můstku k celkové volné energii složeného stavu proteinu provedením bodové mutace, změnou a následným přerušením solného můstku. Například v lysozymu T4 byl identifikován solný můstek mezi kyselinou asparagovou (Asp) na zbytku 70 a histidinem (His) na zbytku 31 (obrázek 3). Byla provedena místně řízená mutageneze s asparaginem (Asn) (obrázek 4), čímž byly získány tři nové mutanty: Asp70Asn His31 (mutant 1), Asp70 His31Asn (mutant 2) a Asp70Asn His31Asn (dvojitý mutant).

Po stanovení mutantů lze k výpočtu volné energie spojené se solným můstkem použít dvě metody. Jedna metoda zahrnuje pozorování teploty tání proteinu divokého typu ve srovnání s teplotou tání tří mutantů. Denaturaci lze sledovat pomocí změny kruhového dichroismu. Snížení teploty tání znamená snížení stability. To se kvantifikuje metodou popsanou Becktelem a Schellmanem, kdy se rozdíl volné energie mezi oběma látkami vypočítá pomocí ΔTΔS. S tímto výpočtem jsou spojeny určité problémy a lze jej použít pouze s velmi přesnými údaji. V příkladu s lysozymem T4 byla ΔS pseudo-wildova typu dříve uváděna při pH 5,5, takže teplotní rozdíl ve středním bodě 11 °C při tomto pH vynásobený uváděnou hodnotou ΔS 360 cal/(mol-K) (1,5 kJ/(mol-K)) dává změnu volné energie přibližně -4 kcal/mol (-17 kJ/mol). Tato hodnota odpovídá množství volné energie, kterou ke stabilitě proteinu přispívá solný můstek.
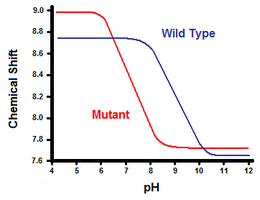
Druhá metoda využívá k výpočtu volné energie solného můstku spektroskopii nukleární magnetické rezonance. Provádí se titrace, přičemž se zaznamenává chemický posun odpovídající protonům uhlíku sousedícího s karboxylátovou nebo amonnou skupinou. Střední bod titrační křivky odpovídá pKa neboli pH, při kterém je poměr protonovaných:deprotonovaných molekul 1:1. Pokračujeme-li v příkladu lysozymu T4, získáme titrační křivku pozorováním posunu protonu C2 histidinu 31 (obrázek 5). Obrázek 5 ukazuje posun titrační křivky mezi divokým typem a mutantem, ve kterém je Asp70 Asn. Solný můstek vzniká mezi deprotonovaným Asp70 a protonovaným His31. Tato interakce způsobuje posun pKa His31. Uvádí se, že v rozloženém proteinu divokého typu, kde solný můstek chybí, má His31 pKa 6,8 v pufrech H20 o střední iontové síle. Obrázek 5 ukazuje pKa divokého typu 9,05. Tento rozdíl v pKa je podpořen interakcí His31 s Asp70. Aby His31 udržel solný můstek, bude se snažit udržet svůj proton co nejdéle. Při narušení solného můstku, jako je tomu u mutanta D70N, se pKa posune zpět na hodnotu 6,9, která je mnohem bližší hodnotě His31 v rozloženém stavu.
Rozdíl v pKa lze kvantifikovat tak, aby odrážel příspěvek solného můstku k volné energii. Pomocí Gibbsovy volné energie:ΔG = -RT ln(Keq), kde R je univerzální plynová konstanta, T je teplota v kelvinech a Keq je rovnovážná konstanta reakce v rovnováze. Deprotonace His31 je kyselá rovnovážná reakce se zvláštní Keq známou jako disociační konstanta kyseliny, Ka: His31-H+ ⇌ His31 + H+. PKa pak souvisí s Ka podle následujícího vzorce: pKa = -log(Ka). Výpočet rozdílu volné energie mutanta a divokého typu lze nyní provést pomocí rovnice volné energie, definice pKa, pozorovaných hodnot pKa a vztahu mezi přirozenými logaritmy a logaritmy. V příkladu lysozymu T4 tento přístup poskytl vypočtený příspěvek k celkové volné energii přibližně 3 kcal/mol. Podobný přístup lze uplatnit u druhého účastníka solného můstku, jako je Asp70 v příkladu s lysozymem T4, sledováním jeho posunu pKa po mutaci His31.
Pozornost při výběru vhodného experimentu se týká umístění solného můstku v proteinu. Prostředí hraje v interakci velkou roli. Při vysokých iontových silách může být solný můstek zcela maskován, protože se jedná o elektrostatickou interakci. Solný můstek His31-Asp70 v lysozymu T4 byl pohřben uvnitř proteinu. Entropie hraje větší roli u povrchových solných můstků, kde jsou zbytky, které mají za normálních okolností možnost pohybu, zúženy jejich elektrostatickou interakcí a vodíkovou vazbou. Ukázalo se, že to snižuje entropii natolik, že se příspěvek interakce téměř vymaže. Povrchové solné můstky lze studovat podobně jako pohřbené solné můstky za použití dvojitých mutantních cyklů a NMR titrací. Ačkoli existují případy, kdy pohřbené solné můstky přispívají ke stabilitě, stejně jako v jiných případech existují výjimky a pohřbené solné můstky mohou vykazovat destabilizující účinek. Rovněž povrchové solné můstky mohou za určitých podmínek vykazovat stabilizační účinek. Stabilizační nebo destabilizační účinek je třeba posuzovat případ od případu a jen málokteré tvrzení je možné učinit paušálně
.