
Isolierung und Charakterisierung von Pterosinderivaten aus P. aquilinum
Pterosinderivate wurden aus den ganzen Pflanzen von P. aquilinum isoliert. Für die Untersuchung der phytochemischen Bestandteile aus dem Rainfarn wurde ein Wasserextrakt nacheinander mit Ethylacetat (EtOAc) und n-Butanol (n-BuOH) partitioniert (Abb. 1). Die wiederholte Säulenchromatographie der in EtOAc löslichen Fraktion führte zur Isolierung von neun Derivaten, darunter (2R)-Pterosin B (1), Pterosin Z (2), (2S)-Pterosin P (3), (3R)-Pterosin D (4), (2S)-Pterosin A (5), (2S,3R)-Pterosin C (6), (2R,3R)-Pterosin C (7), (2R)-Pterosid B (8) und Pterosid Z (9), mit Reinheiten von mehr als 97 % (Abb. 2). 2). Die wiederholte säulenchromatographische Trennung der n-BuOH-löslichen Fraktion führte zur Isolierung von sechs Derivaten, darunter (2S)-Pterosid A (10), (2S)-Pterosid A2 (11), (2S,3R)-Pterosid C (12), (2R,3R)-Pterosid C (13), (3S)-Pterosid D (14) und (2S)-Pterosid P (15), mit Reinheiten von mehr als 97%. Die Strukturen der Verbindungen wurden durch die Analyse der Spektraldaten, einschließlich MS, 1D- und 2D-NMR, identifiziert (Supplementary Information 1).

Strukturen der Pterosinverbindungen 1-15xxx
Hemmende Aktivität der Pterosinderivate gegen BACE1, AChE, und BChE
Um das Anti-AD-Potenzial zu bewerten, wurde die hemmende Wirkung der einzelnen Pterosin-Verbindungen gegen BACE1 und Cholinesterasen durch entsprechende In-vitro-Hemmungsversuche untersucht (Tabelle 1). Alle getesteten Pterosinderivate zeigten konzentrationsabhängige hemmende Aktivitäten gegen BACE1 mit einem Bereich von IC50-Werten (halbmaximale Hemmkonzentration) von 9,74-94,4 μM, mit Ausnahme von (2S)-Pterosin A und (2S)-Pterosid P, die bei den getesteten Konzentrationen inaktiv waren, verglichen mit der IC50 von Quercetin, das als Positivkontrolle verwendet wurde, die 18,8 μM betrug. Die hemmende Wirkung der stärksten Hemmstoffe war in der Reihenfolge (2R,3R)-Pterosid C, (3S)-Pterosid D, (2R)-Pterosid B, (2S,3R)-Pterosin C, (2R,3R)-Pterosin C, (2S,3R)-Pterosid C und (2R)-Pterosin B mit IC50-Werten von 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 bzw. 29,6 μM. Anschließend testeten wir das hemmende Potenzial der Pterosin-Derivate gegen AChE. Alle getesteten Verbindungen zeigten signifikante AChE-hemmende Aktivitäten mit IC50-Werten im Bereich von 2,55-110 μM, verglichen mit dem IC50-Wert gegen AChE von Berberin, das als positive Kontrolle verwendet wurde und 0,39 μM betrug. Die Pterosinverbindungen, die die stärkste hemmende Wirkung gegen AChE zeigten, waren (2R)-Pterosid B, (2R,3R)-Pterosid C, (2S,3R)-Pterosid C, (2S,3R)-Pterosin C und (2R)-Pterosin B mit IC50-Werten von 2,55, 3,77, 9,17, 12,8 bzw. 16,2 μM. Schließlich testeten wir die hemmende Wirkung der Pterosin-Derivate gegen BChE. Alle getesteten Pterosinverbindungen zeigten eine hemmende Wirkung auf BChE mit IC50-Werten zwischen 5,29 und 119 μM, mit Ausnahme von (3R)-Pterosin D, das bei den getesteten Konzentrationen inaktiv war, verglichen mit der IC50 von Berberin gegen BChE, die 3,32 μM betrug. Die Pterosinverbindungen, die die stärkste hemmende Wirkung gegen BChE zeigten, waren (2R,3R)-Pterosid C und Pterosid Z mit IC50-Werten von 5,29 bzw. 5,31 μM.
Gesamt zeigten die meisten der getesteten Pterosinderivate signifikante hemmende Aktivitäten gegen BACE1, AChE und BChE gleichzeitig. Das Vorhandensein der zusätzlichen 2-Hydroxymethyl-Tetrahydro-Pyran-3,4,5-Triol-Gruppe wie in den Pterosid-Derivaten erhöhte die Hemmwirkung gegen die Enzyme erheblich. Außerdem verringerte das Vorhandensein der zusätzlichen Hydroxymethylgruppe an Position 2 des Indanonrings von (2R)-Pterosin B wie in (2S)-Pterosin A oder der Methylgruppe wie in (3R)-Pterosin D und Pterosin Z die hemmenden Aktivitäten gegen die Enzyme. Insbesondere das Vorhandensein der Hydroxymethylgruppe an Position 5 des Indanonrings wie in (2S)-Pterosin P verringerte die hemmende Aktivität gegen BACE1.
Kinetische Parameter der Enzymhemmung durch Pterosinderivate
In einem Versuch, die Art der enzymatischen Hemmung von Pterosinderivaten zu erklären, führten wir eine kinetische Analyse von BACE1 und Cholinesterasen für repräsentative Inhibitoren durch (Tabelle 2, ergänzende Informationen 2). Ein niedriger Ki-Wert (Inhibitionskonstante) deutet auf eine engere Enzymbindung und einen wirksameren Inhibitor hin. Insgesamt korrelierten die Ki-Werte der Verbindungen mit den jeweiligen IC50-Werten. Die BACE1-Hemmung durch die Verbindungen (2R,3R)-Pterosid C, (3S)-Pterosid D und (2R,3R)-Pterosin C war ein Mischtyp mit Ki-Werten von 12,6, 16,5 bzw. 27.6 µM, während die Hemmung durch (2R)-Pterosid B, (2S,3R)-Pterosin C und (2R)-Pterosin B mit Ki-Werten von 23,1, 33,8 bzw. 38,3 µM nichtkompetitiv war. Die AChE-Hemmung durch (2R)-Pterosid B, (2R,3R)-Pterosid C, (2R)-Pterosin B, (2S,3R)-Pterosin C und (3S)-Pterosid D war vom Mischtyp mit Ki-Werten von 4.89, 8,13, 12,1, 16,3 bzw. 23,1 µM, während (2R,3R)-Pterosin C ein nicht-kompetitiver Hemmstoff mit einem Ki-Wert von 29,6 µM war. Die BChE-Hemmung durch (2R,3R)-Pterosin C, (2R,3R)-Pterosid C, (3S)-Pterosid D und (2R)-Pterosid B war ein Mischtyp mit Ki-Werten von 4,77, 9,62, 19.7 bzw. 22,6 µM, während (2S,3R)-Pterosin C und (2R)-Pterosin B nicht-kompetitive Hemmstoffe mit Ki-Werten von 29,9 bzw. 53,5 µM waren. Diese Ergebnisse deuten darauf hin, dass spezifische Pterosin-Derivate wirksame BACE1-, AChE- und BChE-Inhibitoren sein könnten.
Molekulare Docking-Simulationen für BACE1, AChE und BChE
Für BACE1 und Cholinesterasen sind mehrere Kristallstrukturen verfügbar. Wir haben menschliche PDBs auf der Grundlage von Wildtyp-Strukturen, kokristallisierten Liganden und Auflösungen der Strukturen ausgewählt. Die Röntgenkristallstrukturen von BACE1 im Komplex mit QUD (PDB-Code: 2WJO, Auflösung: 2,5 Å)33, AChE im Komplex mit E2020 (PDB-Code: 4EY7, Auflösung: 2,35 Å)34 und BChE im Komplex mit 3F9 (PDB-Code: 4TPK, Auflösung: 2,70 Å)35 wurden für das Docking ausgewählt. Zunächst wurden QUD, E2020 und 3F9 aus den Kristallstrukturen extrahiert und an die aktiven Stellen von BACE1, AChE bzw. BChE angedockt. Anschließend wurden (2R,3R)-Pterosid C, (3S)-Pterosid D, (2R,3R)-Pterosin C, (2R)-Pterosid B, (2S,3R)-Pterosin C und (2R)-Pterosin B mit dem bekannten Hemmungsmechanismus gegen BACE1, AChE und BChE angedockt, um deren SAR zu bestimmen. Die Docking-Ergebnisse sind in Tabelle 3 zusammengefasst. Die SAR der ausgewählten gemischt-kompetitiven und nicht-kompetitiven BACE1-, AChE- und BChE-Inhibitoren ermöglichte die Bewertung der neuartigen, auf die Bindungsstelle gerichteten Hemmung der Enzyme durch Pterosine.
Unser Docking-Modus von E2020 stimmte mit dem experimentell ermittelten Bindungsmodus überein, der zuvor mit rekombinantem menschlichem AChE (rhAChE) berichtet wurde (ergänzende Informationen 3)34. Die quadratische Abweichung (root-mean-square deviation, RMSD) zwischen dem Kristall und den angedockten Konformationen von E2020 betrug 0,54 Å, was auf die Zuverlässigkeit unseres Docking-Setups bei der Reproduktion des experimentellen Bindungsmodus hindeutet. Darüber hinaus führte der angedockte Modus von E2020 zu einer ähnlichen Interaktion wie der von rhAChE-E2020. In unserer Studie wurden die Wassermoleküle während des Andockens aus der Kristallstruktur entfernt; daher wurden die durch Wasser vermittelten Wechselwirkungen in der vorliegenden Studie nicht analysiert. In ähnlicher Weise stimmten die angedockten Modi von QUD und 3F9 mit den verfügbaren experimentellen Daten für BACE1 33 bzw. BChE35 überein (ergänzende Informationen 3). Die RMSDs zwischen den Kristall- und angedockten Konformationen von QUD und 3F9 betrugen 0,46 bzw. 0,60 Å. Außerdem stimmten die Bindungsstellen der Pterosin-Inhibitoren mit einer früheren Docking-Studie überein, bei der BACE1, AChE und BChE38 beteiligt waren. In der Studie wurde jedoch Tetronarce californica AChE (PDB-Code: 1ACJ) verwendet, das aufgrund von Variationen in ihren Sequenzen etwas andere Restnummern als menschliches AChE enthält.
BACE1-Docking
Auf der Grundlage des Hemmungstyps und der Aktivität wurden (2R,3R)-Pterosid C und (2R)-Pterosid B ausgewählt, um die angedockten Modi von BACE1-Hemmern des gemischten Typs bzw. des nichtkompetitiven Typs zu zeigen. Abbildung 3a und b zeigen die Docking-Modelle von (2R,3R)-Pterosid C bzw. (2R)-Pterosid B. Die Interaktionen der angedockten Verbindungen innerhalb des aktiven Zentrums von BACE1 sind in Abb. 4 dargestellt.

Molekulare Docking-Modelle für a den BACE1-Inhibitor vom Mischtyp (2R,3R)-Pterosid C (blaue Farbe) und b den nichtkompetitiven BACE1-Inhibitor (2R)-Pterosid B (braune Farbe). Die angedockten Posen sind der Röntgenkristallstruktur von QUD (gelbe Farbe) überlagert (PDB-Code: 2WJO). BACE1, die Reste des aktiven Zentrums und die Verbindungen sind durch Band-, Linien- bzw. Stabmodelle dargestellt. Die Farben der gepunkteten Linien erklären die Art der verschiedenen Wechselwirkungen: Wasserstoffbrückenbindungen (grün), hydrophobe Wechselwirkungen (rosa) und π-Sigma-Wechselwirkungen (lila). BACE1 β-site amyloid precursor protein cleaving enzyme 1

Ligandeninteraktionsdiagramm von a QUD, b (2R,3R)-Pterosid C, c (3S)-Pterosid D, d (2R,3R)-Pterosin C, e (2R)-Pterosid B, f (2S,3R)-Pterosin C und g (2R)-Pterosin B im aktiven Zentrum von BACE1. Die Farben der gepunkteten Linien erklären die Art der verschiedenen Wechselwirkungen: Wasserstoffbrückenbindungen (grün), hydrophobe Wechselwirkungen (rosa) und π-Sigma-Wechselwirkungen (lila). BACE1 β-site amyloid precursor protein cleaving enzyme 1
Die angedockte Pose von QUD wies eine Bindungsenergie (B.E.) von -7,59 kcal/mol auf. Wie in Abb. 4a gezeigt, wies die NH2-Gruppe am Chinazolinring des Liganden zwei Wasserstoffbrückenbindungen mit den CO-Gruppen von Asp32 und Asp228 in Abständen von 1,86 bzw. 2,17 Å auf. Eine dritte Wasserstoffbrücke wurde zwischen der anderen NH-Gruppe des Liganden und der CO-Gruppe von Gly230 in einem Abstand von 2,16 Å beobachtet. Leu30, Tyr71, Phe108 und Val332 vermittelten die hydrophoben Wechselwirkungen. Abbildung 4b-d zeigt die angedockten Posen von (2R,3R)-Pterosid C, (3S)-Pterosid D und (2R,3R)-Pterosin C (BACE1-Inhibitoren vom Mischtyp). Sie wurden in der Bindungstasche positioniert, die von Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 und Thr231 begrenzt wird. Hinsichtlich ihrer Aktivität wiesen (2R,3R)-Pterosid C (IC50 = 9,74 µM), (3S)-Pterosid D (IC50 = 10,7 µM) und (2R,3R)-Pterosin C (IC50 = 26,2 µM) einen B.E. von -7,27, -6,93 bzw. -4,84 kcal/mol auf. (2R,3R)-Pterosid C wies eine höhere Wirksamkeit als (2R,3R)-Pterosin C auf, was auf das Vorhandensein einer zusätzlichen 2-Hydroxymethyl-tetrahydro-pyran-3,4,5-triol-Gruppe zurückzuführen war, die drei Wasserstoffbrückenbindungen bildete (Abb. 4b). Die OH-Gruppe der Hydroxymethyl- und die 3-OH-Gruppe des Tetrahydro-Pyran-Triol-Rings zeigten zwei Wasserstoffbrückenbindungen mit der CO-Gruppe von Asp228 in Abständen von 2,63 bzw. 2,12 Å. Zusätzlich zeigte die 3-OH-Gruppe eine Wasserstoffbrückenbindung mit Thr231 in einem Abstand von 1,89 Å. Im Fall von (3S)-Pterosid D (Abb. 4c) veränderte das Vorhandensein der 2,2-Dimethylgruppe am Indanonring die Bindungsinteraktionen leicht im Vergleich zu (2R,3R)-Pterosid C. Die 3-OH-Gruppe des Tetrahydro-Pyran-Triol-Rings zeigte nur eine einzige Wasserstoffbrückenbindung mit Asp228 in einem Abstand von 1,93 Å. Die 3-OH-Gruppe des Indanonrings wies jedoch eine zusätzliche Wasserstoffbrückenbindung mit Ile126 in einem Abstand von 2,34 Å auf. Diese Wechselwirkungen verringerten die Aktivität von (3S)-Pterosid D im Vergleich zu (2R,3R)-Pterosid C leicht. Die in Abb. 4d dargestellten Docking-Interaktionen von (2R,3R)-Pterosin C zeigen, dass die 3-OH-Gruppe des Indanonrings über zwei Wasserstoffbrückenbindungen in Abständen von 1,83 bzw. 2,07 Å an Ser36 und Asn37 gebunden ist. Ala39, Val69, Trp76, Ile118 und Arg128 waren an hydrophoben Wechselwirkungen beteiligt, während Val69 eine π-Sigma-Wechselwirkung zeigte.
Die angedockten Posen von (2R)-Pterosid B, (2S,3R)-Pterosin C und (2R)-Pterosin B (nichtkompetitive BACE1-Inhibitoren) sind in Abb. 4e-g dargestellt. Sie wurden an den von Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 und Ile126 umschlossenen Hohlraum angedockt. In Übereinstimmung mit ihren Aktivitätswerten wiesen (2R)-Pterosid B (IC50 = 18,0 µM), (2S,3R)-Pterosin C (IC50 = 23,1 µM) und (2R)-Pterosin B (IC50 = 29,6 µM) einen B.E. von -6,16, -5,07 bzw. -4,64 kcal/mol auf. Wie in Abb. 4e dargestellt, zeigte (2R)-Pterosid B eine höhere Aktivität als (2S,3R)-Pterosin C und (2R)-Pterosin B, was auf das Vorhandensein einer zusätzlichen 2-Hydroxymethyl-tetrahydro-pyran-3,4,5-triol-Gruppe zurückzuführen ist, die vier Wasserstoffbrückenbindungen aufweist. Zwei Wasserstoffbrückenbindungen wurden zwischen der 4-OH-Gruppe des Tetrahydro-pyran-triol-Rings und den NH- und CO-Gruppen von Asn37 in Abständen von 2,48 bzw. 2,19 Å beobachtet. Darüber hinaus wiesen die 3- und 5-OH-Gruppen zwei zusätzliche Wasserstoffbrückenbindungen mit Ile126 und Trp76 in Abständen von 2,19 bzw. 2,46 Å auf. (2S,3R)-Pterosin C zeigte eine etwas bessere Aktivität als (2R)-Pterosin B, was auf das Vorhandensein einer zusätzlichen OH-Gruppe an Position 3 des Indanonrings zurückzuführen ist, die eine Wasserstoffbrücke mit Lys107 in einem Abstand von 2,12 Å bildete (Abb. 4f). Die anderen Wechselwirkungen waren denen von (2R)-Pterosin B ähnlich. Wie in Abb. 4g dargestellt, zeigte (2R)-Pterosin B hydrophobe Wechselwirkungen mit Val69, Tyr76 und Phe108.
AChE-Docking
(2R)-Pterosid B und (2R,3R)-Pterosin C wurden als Vertreter ausgewählt, um die angedockten Modi von AChE-Inhibitoren des gemischten Typs bzw. des nicht-kompetitiven Typs aufgrund ihrer Aktivitäten und der Art ihrer AChE-Hemmung zu demonstrieren. Abbildung 5a und b zeigen die Docking-Modelle von (2R)-Pterosid B bzw. (2R,3R)-Pterosin C. Die Wechselwirkungen der angedockten Verbindungen innerhalb des aktiven Zentrums von AChE sind in Abb. 6 dargestellt.

Molekulare Docking-Modelle für a den gemischten AChE-Inhibitor (2R)-Pterosid B (blaue Farbe) und b den nichtkompetitiven AChE-Inhibitor (2R,3R)-Pterosin C (braune Farbe). Die angedockten Posen sind der Röntgenkristallstruktur von E2020 (gelbe Farbe) überlagert (PDB-Code: 4EY7). AChE, die Reste des aktiven Zentrums und die Verbindungen sind durch Band-, Linien- bzw. Stabmodelle dargestellt. Die Farben der gepunkteten Linien erklären die Art der verschiedenen Wechselwirkungen: Wasserstoffbrückenbindungen (grün) und hydrophobe Wechselwirkungen (rosa). AChE Acetylcholinesterase

Ligandeninteraktionsdiagramm von a E2020, b (2R)-Pterosid B, c (2R,3R)-Pterosid C, d (2S,3R)-Pterosin C, e (2R)-Pterosin B, f (3S)-Pterosid D und g (2R,3R)-Pterosin C im aktiven Zentrum von AChE. Die Farben der gepunkteten Linien erklären die Art der verschiedenen Wechselwirkungen: Wasserstoffbrückenbindungen (grün), hydrophobe Wechselwirkungen (rosa) und π-Sigma-Wechselwirkungen (lila). AChE Acetylcholinesterase
Die angedockte Pose von E2020 zeigte einen B.E. von -10,28 kcal/mol. Wie in Abb. 6a dargestellt, bildete die CO-Gruppe des Indanonrings eine Wasserstoffbrücke mit der NH-Gruppe von Phe295 in einem Abstand von 1,70 Å. Trp286 und Tyr341 waren an π-Sigma-Wechselwirkungen beteiligt, während Trp86, Trp286, Tyr337, Phe338 und Tyr341 hydrophobe Wechselwirkungen vermittelten. Abbildung 6b-f zeigt die angedockten Posen von (2R)-Pterosid B, (2R,3R)-Pterosid C, (2S,3R)-Pterosin C, (2R)-Pterosin B und (3S)-Pterosid D (AChE-Inhibitoren vom Mischtyp). Sie befanden sich im aktiven Zentrum, umgeben von Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 und Tyr341. In Übereinstimmung mit ihren Aktivitätswerten wurden (2R)-Pterosid B (IC50 = 2,55 µM), (2R,3R)-Pterosid C (IC50 = 3,77 µM), (2S,3R)-Pterosin C (IC50 = 12.8 µM), (2R)-Pterosin B (IC50 = 16,2 µM) und (3S)-Pterosid D (IC50 = 27,4 µM) wiesen einen B.E. von -7,90, -7,49, -6,03, -5,76 bzw. -4,91 kcal/mol auf. (2R)-Pterosid B zeigte eine höhere Potenz als (2S,3R)-Pterosin C und (2R)-Pterosin B, was auf das Vorhandensein einer zusätzlichen 2-Hydroxymethyl-tetrahydro-pyran-3,4,5-triol-Gruppe zurückzuführen ist, die drei Wasserstoffbrückenbindungen herstellt (Abb. 6b). Die OH-Gruppe der Hydroxymethyl- und die 3-OH-Gruppe des Tetrahydro-Pyran-Triol-Rings zeigten Wasserstoffbrückenbindungen mit der CO-Gruppe von Asn87 in Abständen von 2,23 bzw. 2,28 Å. Außerdem bildete die 4-OH-Gruppe eine Wasserstoffbrücke mit der CO-Gruppe von Trp86 in einem Abstand von 2,13 Å. Im Fall von (2R,3R)-Pterosid C (Abb. 6c) zeigte die Methylgruppe an Position-2 des Indanonrings keine hydrophobe Wechselwirkung mit Tyr341 und wies daher eine vergleichsweise geringere Aktivität auf als (2R)-Pterosid B. Tyr341 behielt jedoch die hydrophobe Wechselwirkung mit dem anderen Teil des Indanonrings bei, wie in (2R)-Pterosid B gezeigt. Die höhere Aktivität von (2S,3R)-Pterosin C im Vergleich zu (2R)-Pterosin B wurde auf das Vorhandensein einer zusätzlichen OH-Gruppe an Position-3 des Indanonrings zurückgeführt, die eine Wasserstoffbrücke mit Phe295 in einem Abstand von 1,82 Å bildete (Abb. 6d). Die übrigen Wechselwirkungen waren mit den (2R)-Pterosin B-Wechselwirkungen vergleichbar. Wie in Abb. 6e gezeigt, bildete die OH-Gruppe der Hydroxyethylgruppe an Position 6 des Indanonrings eine Wasserstoffbrücke mit Tyr124 in einem Abstand von 2,48 Å. Tyr124, Trp286, Tyr337, Phe338 und Tyr341 trugen zu den hydrophoben Wechselwirkungen bei. Im Vergleich zu (2R)-Pterosid B und (2R,3R)-Pterosid C zeigte (3S)-Pterosid D aufgrund des Vorhandenseins der 2,2-Dimethylgruppe am Indanonring unähnliche Bindungsinteraktionen (Abb. 6f). Die 2,2-Dimethylgruppe trug wesentlich zu der ausgeprägten Andockstellung von (3S)-Pterosid D bei. Die 5-OH-Gruppe des Tetrahydro-Pyran-Triol-Rings bildete eine Wasserstoffbrückenbindung mit Asp74 in einem Abstand von 1,96 Å. Die CO-Gruppe des Indanonrings zeigte eine Wasserstoffbrückenbindung mit Tyr72 in einem Abstand von 2,97 Å. Diese Wechselwirkungen waren für die geringe Aktivität von (3S)-Pterosid D verantwortlich.
Die angedockte Position von (2R,3R)-Pterosin C (nichtkompetitiver AChE-Inhibitor) ist in Abb. 6g dargestellt. (2R,3R)-Pterosin C (IC50 = 23,2 µM) zeigte einen B.E. von -5,01 kcal/mol. Die Bindungstasche von (2R,3R)-Pterosin C umfasste Tyr72, Trp286, Ser293, Phe297 und Tyr341, mit zwei Wasserstoffbrückenbindungen. Eine der Wasserstoffbrücken wurde zwischen der 3-OH-Gruppe des Indanonrings und der CO-Gruppe von Ser293 in einem Abstand von 2,03 Å gebildet. Die zweite Wasserstoffbrücke wurde zwischen der OH-Gruppe der Hydroxyethylgruppe an Position 6 des Indanonrings und Tyr72 in einem Abstand von 1,94 Å beobachtet. Reste wie Tyr72, Trp286, Phe297 und Tyr341 nahmen an hydrophoben Wechselwirkungen teil.
BChE-Docking
Unter Berücksichtigung der Aktivitätsniveaus und der Art der BChE-Hemmung wurden (2R,3R)-Pterosid C und (2S,3R)-Pterosin C ausgewählt, um die angedockten Modi von BChE-Hemmern des gemischten Typs bzw. des nichtkompetitiven Typs zu demonstrieren. Abbildung 7a, b zeigt die Docking-Modelle von (2R,3R)-Pterosid C und (2S,3R)-Pterosin C. Die Wechselwirkungen der angedockten Verbindungen innerhalb des BChE-Aktivzentrums sind in Abb. 8 dargestellt.
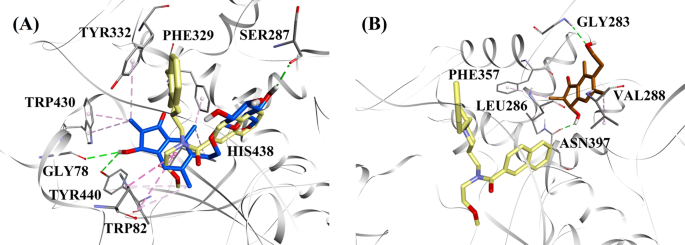
Molekulare Docking-Modelle für a den BChE-Hemmer vom Mischtyp (2R,3R)-Pterosid C (blaue Farbe) und b den nichtkompetitiven BChE-Hemmer (2S,3R)-Pterosin C (braune Farbe). Die angedockten Posen sind der Röntgenkristallstruktur von 3F9 (gelbe Farbe) überlagert (PDB-Code: 4TPK). BChE, die Reste des aktiven Zentrums und die Verbindungen sind durch Band-, Linien- bzw. Stabmodelle dargestellt. Die Farben der gepunkteten Linien erklären die Art der verschiedenen Wechselwirkungen: Wasserstoffbrückenbindungen (grün) und hydrophobe Wechselwirkungen (rosa). BChE Butyrylcholinesterase
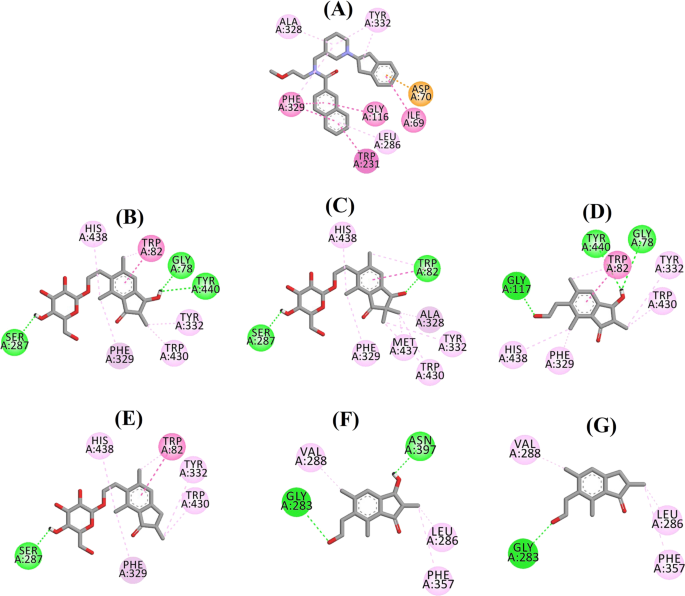
Ligandeninteraktionsdiagramm von a 3F9, b (2R,3R)-Pterosid C, c (3S)-Pterosid D, d (2R,3R)-Pterosin C, e (2R)-Pterosid B, f (2S,3R)-Pterosin C und g (2R)-Pterosin B im aktiven Zentrum von BChE. Die Farben der gepunkteten Linien erklären die Art der verschiedenen Wechselwirkungen: Wasserstoffbrückenbindungen (grün), hydrophobe Wechselwirkungen (rosa) und π-Anionen-Wechselwirkungen (golden). BChE Butyrylcholinesterase
Die angedockte Pose von 3F9 zeigte einen B.E. von -8,49 kcal/mol. Wie in Abb. 8a dargestellt, waren hauptsächlich hydrophobe Wechselwirkungen für die Ligandenbindung verantwortlich. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 und Tyr332 waren für die hydrophoben Wechselwirkungen verantwortlich, während Asp70 eine π-Anionenwechselwirkung zeigte. Abbildung 8b-e zeigt die angedockten Posen von (2R,3R)-Pterosid C, (3S)-Pterosid D, (2R,3R)-Pterosin C und (2R)-Pterosid B (BChE-Hemmer vom Mischtyp). Ihre Bindungstasche bestand aus Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 und Tyr440. Entsprechend ihrer Aktivität wiesen (2R,3R)-Pterosid C (IC50 = 5,29 µM), (3S)-Pterosid D (IC50 = 19,3 µM), (2R,3R)-Pterosin C (IC50 = 20,3 µM) und (2R)-Pterosid B (IC50 = 62,0 µM) einen B.E. von -7,23, -6,59, -6,52 bzw. -4,38 kcal/mol auf. Wie in Abb. 8b zu sehen ist, wies die 3-OH-Gruppe des Indanonrings von (2R,3R)-Pterosid C zwei Wasserstoffbrückenbindungen mit Gly78 und Tyr440 bei Abständen von 2,87 bzw. 2,89 Å auf. Im Fall von (3S)-Pterosid D beeinflusste die 2,2-Dimethylgruppe am Indanonring die Bindungswechselwirkungen der 3-OH-Gruppe (Abb. 8c). Die 3-OH-Gruppe bildete nur eine Wasserstoffbrückenbindung mit Trp82 in einem Abstand von 2,94 Å, was zu einer vergleichsweise geringeren Aktivität von (3S)-Pterosid D als (2R,3R)-Pterosid C führte. Wie in Abb. 8d zu sehen ist, konnte (2R,3R)-Pterosin C aufgrund des Fehlens einer 2-Hydroxymethyl-tetrahydro-pyran-3,4,5-triol-Gruppe keine Wasserstoffbrückenbindung mit Ser287 herstellen. Folglich war seine Aktivität geringer als die von (2R,3R)-Pterosid C. Das Fehlen der OH-Gruppe an Position 3 des Indanonrings war für die sehr geringe Aktivität von (2R)-Pterosid B verantwortlich (Abb. 8e). 8e), das ähnlich wie (2R,3R)-Pterosid C keine Wasserstoffbrückenbindungen mit Gly78 und Tyr440 bilden konnte.
Die angedockten Posen von (2S,3R)-Pterosin C und (2R)-Pterosin B (nichtkompetitive BChE-Inhibitoren) sind in Abb. 8f bzw. g dargestellt. Diese angedockten Posen befanden sich in dem von Gly283, Leu286, Val288, Phe357 und Asn397 umschlossenen Hohlraum. Entsprechend ihrer Aktivität wiesen (2S,3R)-Pterosin C (IC50 = 44,3 µM) und (2R)-Pterosin B (IC50 = 48,1 µM) einen B.E. von -5,40 bzw. -5,06 kcal/mol auf. (2S,3 R)-Pterosin C war wirksamer als (2R)-Pterosin B aufgrund des Vorhandenseins einer zusätzlichen OH-Gruppe an Position 3 des Indanonrings, die eine Wasserstoffbrücke mit der CO-Gruppe von Asn397 in einem Abstand von 2,05 Å bildete (Abb. 8f). Andere Wechselwirkungen waren ähnlich wie bei (2R)-Pterosin B. Wie in Abb. 8g gezeigt, bildete die OH-Gruppe in der Hydroxyethylgruppe an Position-6 des Indanonrings eine Wasserstoffbrücke mit Gly283 in einem Abstand von 2,40 Å. Die Reste Leu286, Val288 und Phe357 waren an hydrophoben Wechselwirkungen beteiligt.
Inhibitoren vom Mischtyp binden sowohl an das freie Enzym als auch an den Enzym-Substrat-Komplex, was darauf hindeutet, dass diese Verbindungen an die katalytische Stelle des jeweiligen Enzyms binden können. Nichtkompetitive Inhibitoren binden an die allosterische Stelle des freien Enzyms oder des Enzym-Substrat-Komplexes. Eine kürzlich durchgeführte Studie legt nahe, dass kompetitive, gemischte und nichtkompetitive Inhibitoren unterschiedliche Stellen in den Bindungstaschen von BACE1, AChE und BChE38 besetzen. Beim Docking zur Bewertung des hemmenden Mechanismus von Pterosin-Derivaten wurden die Bindungsstellen der Verbindungen entsprechend ihrer Art der Hemmung definiert. Die Docking-Ergebnisse zeigten, dass sich die Bindungsstellen von gemischten und nichtkompetitiven Inhibitoren für BACE1, AChE und BChE an den entsprechenden aktiven Stellen teilweise überlappen und mit einem früheren Bericht übereinstimmen38.
BBB-Permeabilität
PAMPA-BBB, ein in vitro-Test für die künstliche Membranpermeabilität der BHS, ist eines der zuverlässigsten physikalisch-chemischen Screening-Instrumente in der frühen Phase der Entdeckung von auf das ZNS abzielenden Arzneimitteln40. Das PAMPA-BBB-System modelliert die transzelluläre passive Diffusion von Chemikalien durch die BHS und misst streng passive Transportmechanismen über eine künstliche Lipidmembran anhand der effektiven Permeabilität (Pe, cm/s). Auf der Grundlage des für die Vorhersage der BHS-Permeation erstellten Musters wurden die Verbindungen in folgende Kategorien eingeteilt: (i) „ZNS+“ (hohe vorhergesagte BHS-Permeation); Pe (10-6 cm/s) > 4,00, (ii) „ZNS-“ (niedrige vorhergesagte BHS-Permeation); Pe (10-6 cm/s) < 2,00 und (iii) „ZNS+/-“ (BHS-Permeation unsicher); Pe (10-6 cm/s) von 4,00 bis 2,00. Dementsprechend zeigten (2R)-Pterosin B, (2S)-Pterosin P und (2S)-Pterosin A eine hohe BBB-Permeation mit Pe-Werten von 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s bzw. 6,26 × 10-6 cm/s (Tabelle 4, ergänzende Informationen 4). Der Pe-Wert von (2R)-Pterosin B war 1,7-mal höher als der des ZNS-Medikaments Verapamil (Pe = 34,6 × 10-6 cm/s), das als Positivkontrolle verwendet wurde. (2S,3R)-Pterosin C und (2R,3R)-Pterosin C zeigten eine unsichere BHS-Permeation mit Pe-Werten von 2,34 bzw. 1,98. (2R,3R)-Pterosid C, (3S)-Pterosid D und (2R)-Pterosid B, die unter den getesteten Pterosin-Derivaten die stärkste BACE1- und Cholinesterase-hemmende Wirkung zeigten, wiesen eine sehr geringe BBB-Permeabilität auf. Das Vorhandensein der 2-Hydroxymethyl-Tetrahydro-Pyran-3,4,5-Triol-Gruppe wie in den Pterosid-Derivaten führte zu einer bemerkenswerten Verringerung der BHS-Permeabilität. Im Vergleich zu (2R)-Pterosin B führte das zusätzliche Vorhandensein der OH-Gruppe an Position-3 des Indanonrings wie in Pterosin C, der Hydroxymethylgruppe an Position-2 des Indanonrings wie in (2S)-Pterosin A oder der Hydroxymethylgruppe an Position-5 des Indanonrings wie in (2S)-Pterosin P ebenfalls zu einer deutlichen Verringerung der BHS-Permeabilität. In Anbetracht einer außergewöhnlich hohen BHS-Permeabilität und der signifikanten Hemmung von BACE1, AChE und BChE könnte (2R)-Pterosin B das Potenzial haben, eine starke Anti-AD-Aktivität zu zeigen.
Effekte von (2R)-Pterosin B und (2R,3R)-Pterosid C auf die Sekretion von Aβ-Peptiden durch neuronale Zellen
Um die Funktion von (2R)-Pterosin B und (2R,3R)-Pterosid C bei der Verringerung der Ausscheidung von Aβ aus neuronalen Zellen zu untersuchen, haben wir eine murine Neuroblastom-Zelllinie verwendet, die das menschliche APPswe stabil überexprimiert. Die Zelllinie ist ein zelluläres Modell der Alzheimer-Krankheit, das durch eine übermäßige Sekretion von Aβ40 und Aβ42 gekennzeichnet ist. Die toxischen Amyloid-Oligomere werden aus den beiden Isoformen des Aβ-Peptids mit unterschiedlichen Längen gebildet. Aβ40 ist die im Gehirn am häufigsten vorkommende Aβ-Isoform41, während Aβ42 bei bestimmten Formen von Alzheimer signifikant zunimmt42. Der Sandwich-ELISA von Aβ40 zeigte, dass (2R)-Pterosin B die Menge an Aβ40-Peptid, die aus den Neuroblastomzellen in das Medium sezerniert wurde, bei 500 μM signifikant um bis zu 50 % reduzierte (P < 0,01) (Abb. 9a). Ebenso nahm die Sekretion von Aβ42-Peptid durch die Neuroblastomzellen in Gegenwart von 500 µM (2R,3R)-Pterosid C signifikant ab (P < 0,05) (Abb. 9b). Zusammenfassend lässt sich sagen, dass (2R)-Pterosin B und (2R,3R)-Pterosid C die Sekretion von Aβ-Peptiden aus Neuroblastomzellen in einer Konzentration von 500 μM signifikant verringerten.

a Wirkung von (2R)-Pterosin B auf die Sekretion von Aβ-Peptiden. APPswe-sezernierende Neuroblastomzellen wurden 24 Stunden lang mit 12, 60, 250 oder 500 µM (2R)-Pterosin B behandelt, und konditionierte Medien wurden in Gegenwart eines Proteaseinhibitors gesammelt. β-SI (10 µM) wurde als positive Kontrolle verwendet. Die Negativkontrolle umfasste Zellen, die in Abwesenheit der Testverbindungen kultiviert wurden. Die quantitative Analyse von sezerniertem Aβ40 und Aβ42 in den konditionierten Medien wurde mittels Sandwich-ELISA durchgeführt. Dargestellt sind die Mittelwerte ± SD von drei unabhängigen Experimenten. Die sezernierten Aβ40-Peptide nahmen in Gegenwart von 500 µM (2R)-Pterosin B signifikant ab. CON Negativkontrolle, β-SI β-Sekretase-Inhibitor III, Aβ β-Amyloid. *P < 0,05, **P < 0,01. b Wirkung von (2R,3R)-Pterosin C auf die Sekretion von Aβ-Peptiden. APPswe-sezernierende Neuroblastomzellen wurden 24 Stunden lang mit 12, 60, 125, 250 oder 500 µM (2R,3R)-Pterosid C behandelt, und konditionierte Medien wurden in Gegenwart eines Proteaseinhibitors gesammelt. β-SI (10 µM) wurde als positive Kontrolle verwendet. Die Negativkontrolle umfasste Zellen, die in Abwesenheit der Testverbindungen kultiviert wurden. Die quantitative Analyse von sezerniertem Aβ40 und Aβ42 in den konditionierten Medien wurde mittels Sandwich-ELISA durchgeführt. Dargestellt sind die Mittelwerte ± SD von drei unabhängigen Experimenten. Das sekretierte Aβ42-Peptid nahm in Gegenwart von 500 µM (2R,3R)-Pterosid C signifikant ab. CON Negativkontrolle, β-SI β-Sekretase-Inhibitor III, Aβ β-Amyloid. *P < 0.05
Zytotoxizität von Pterosin-Derivaten basierend auf MTT-Assay43
Insgesamt zeigten Pterosin-Derivate vernachlässigbare Zytotoxizität gegen verschiedene normale und Krebs-Zelllinien, wie SH-SY5Y menschliches Neuroblastom, C6 Ratten-Gliazellen, NIH3T3 Maus-Embryo-Fibroblasten und B16F10 Maus-Melanom mit LD50-Werten über 0.5 mM (Ergänzende Information 5). Insbesondere (2R)-Pterosin B, (3R)-Pterosin D, (2S)-Pterosin P, (2S)-Pterosid A, (2R)-Pterosid B und (2R,3R)-Pterosid C zeigten keine Zytotoxizität gegen die getesteten Zelllinien mit LD50-Werten über 5 mM. Interessanterweise zeigten mehrere Pterosine relative antiproliferative Wirkungen gegen neuronale SH-SY5Y-Zellen im Vergleich zu C6-Gliazellen mesenchymalen Ursprungs. Die vorliegenden Ergebnisse stimmen mit einem früheren Bericht überein, der darauf hindeutet, dass Pterosinderivate für den Menschen ungiftig sind26.