

Der Beitrag einer Salzbrücke zur Gesamtstabilität des gefalteten Zustands eines Proteins kann durch thermodynamische Daten aus Mutagenesestudien und Kernspinresonanztechniken bewertet werden. Unter Verwendung eines mutierten Pseudo-Wildtyp-Proteins, das speziell mutiert wurde, um eine Ausfällung bei hohem pH-Wert zu verhindern, kann der Beitrag der Salzbrücke zur gesamten freien Energie des gefalteten Proteinzustands bestimmt werden, indem eine Punktmutation durchgeführt wird, die die Salzbrücke verändert und folglich bricht. So wurde beispielsweise im T4-Lysozym eine Salzbrücke zwischen Asparaginsäure (Asp) am Rest 70 und einem Histidin (His) am Rest 31 identifiziert (Abbildung 3). Durch ortsgerichtete Mutagenese mit Asparagin (Asn) (Abbildung 4) wurden drei neue Mutanten erhalten: Asp70Asn His31 (Mutante 1), Asp70 His31Asn (Mutante 2) und Asp70Asn His31Asn (Doppelmutante).

Sobald die Mutanten etabliert sind, können zwei Methoden zur Berechnung der mit einer Salzbrücke verbundenen freien Energie angewendet werden. Eine Methode besteht in der Beobachtung der Schmelztemperatur des Wildtyp-Proteins im Vergleich zu der der drei Mutanten. Die Denaturierung kann durch eine Veränderung des Zirkulardichroismus verfolgt werden. Eine Verringerung der Schmelztemperatur deutet auf eine Verringerung der Stabilität hin. Dies wird durch eine von Becktel und Schellman beschriebene Methode quantifiziert, bei der die Differenz der freien Energie zwischen den beiden durch ΔTΔS berechnet wird. Diese Berechnung ist nicht ganz unproblematisch und kann nur bei sehr genauen Daten angewendet werden. Im Beispiel von T4-Lysozym wurde ΔS des Pseudo-Wildtyps zuvor bei pH 5,5 angegeben, so dass der mittlere Temperaturunterschied von 11 °C bei diesem pH multipliziert mit dem angegebenen ΔS von 360 cal/(mol-K) (1,5 kJ/(mol-K)) eine Änderung der freien Energie von etwa -4 kcal/mol (-17 kJ/mol) ergibt. Dieser Wert entspricht dem Anteil der freien Energie, der durch die Salzbrücke zur Stabilität des Proteins beiträgt.
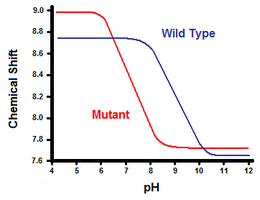
Die zweite Methode nutzt die Kernspinresonanzspektroskopie zur Berechnung der freien Energie der Salzbrücke. Man führt eine Titration durch und zeichnet dabei die chemische Verschiebung auf, die den Protonen des an die Carboxylat- oder Ammoniumgruppe angrenzenden Kohlenstoffs entspricht. Der Mittelpunkt der Titrationskurve entspricht dem pKa oder dem pH-Wert, bei dem das Verhältnis von protonierten zu deprotonierten Molekülen 1:1 beträgt. Am Beispiel von T4-Lysozym lässt sich eine Titrationskurve durch Beobachtung einer Verschiebung des C2-Protons von Histidin 31 ermitteln (Abbildung 5). Abbildung 5 zeigt die Verschiebung der Titrationskurve zwischen dem Wildtyp und der Mutante, bei der Asp70 Asn ist. Die gebildete Salzbrücke besteht zwischen dem deprotonierten Asp70 und dem protonierten His31. Diese Wechselwirkung verursacht die Verschiebung des pKa-Wertes von His31. Im ungefalteten Wildtyp-Protein, bei dem die Salzbrücke nicht vorhanden ist, hat His31 in H20-Puffern mit mäßiger Ionenstärke einen pKa-Wert von 6,8. Abbildung 5 zeigt einen pKa-Wert des Wildtyps von 9,05. Dieser Unterschied im pKa wird durch die Wechselwirkung von His31 mit Asp70 unterstützt. Um die Salzbrücke aufrechtzuerhalten, versucht His31, sein Proton so lange wie möglich zu behalten. Wenn die Salzbrücke unterbrochen wird, wie bei der Mutante D70N, verschiebt sich der pKa auf einen Wert von 6,9 zurück, was dem von His31 im ungefalteten Zustand sehr viel näher kommt.
Der Unterschied im pKa kann quantifiziert werden, um den Beitrag der Salzbrücke zur freien Energie zu ermitteln. Unter Verwendung der freien Energie nach Gibbs:ΔG = -RT ln(Keq), wobei R die universelle Gaskonstante, T die Temperatur in Kelvin und Keq die Gleichgewichtskonstante einer Reaktion im Gleichgewicht ist. Die Deprotonierung von His31 ist eine Säuregleichgewichtsreaktion mit einer speziellen Keq, die als Säuredissoziationskonstante Ka bekannt ist: His31-H+ ⇌ His31 + H+. Der pKa-Wert ist dann mit Ka wie folgt verknüpft: pKa = -log(Ka). Mit Hilfe der Gleichung für die freie Energie, der Definition von pKa, der beobachteten pKa-Werte und der Beziehung zwischen natürlichen Logarithmen und Logarithmen kann nun die Differenz der freien Energie der Mutante und des Wildtyps errechnet werden. Im Beispiel des T4-Lysozyms ergab dieser Ansatz einen berechneten Beitrag von etwa 3 kcal/mol zur gesamten freien Energie. Ein ähnlicher Ansatz kann mit dem anderen Teilnehmer an der Salzbrücke, wie Asp70 im T4-Lysozym-Beispiel, verfolgt werden, indem seine pKa-Verschiebung nach der Mutation von His31 beobachtet wird.
Ein Wort der Vorsicht bei der Wahl des geeigneten Experiments betrifft die Lage der Salzbrücke innerhalb des Proteins. Die Umgebung spielt eine große Rolle bei der Wechselwirkung. Bei hohen Ionenstärken kann die Salzbrücke vollständig verdeckt werden, da es sich um eine elektrostatische Wechselwirkung handelt. Die His31-Asp70-Salzbrücke in T4-Lysozym war innerhalb des Proteins vergraben. Entropie spielt eine größere Rolle bei Oberflächensalzbrücken, bei denen Reste, die sich normalerweise bewegen können, durch ihre elektrostatische Wechselwirkung und Wasserstoffbrückenbindungen eingeengt werden. Es hat sich gezeigt, dass dies die Entropie so weit verringert, dass der Beitrag der Wechselwirkung nahezu aufgehoben wird. Oberflächen-Salzbrücken können ähnlich wie vergrabene Salzbrücken untersucht werden, wobei Doppelmutantenzyklen und NMR-Titrationen eingesetzt werden. Obwohl es Fälle gibt, in denen vergrabene Salzbrücken zur Stabilität beitragen, gibt es wie bei allem anderen auch Ausnahmen, und vergrabene Salzbrücken können eine destabilisierende Wirkung haben. Auch Oberflächen-Salzbrücken können unter bestimmten Bedingungen eine stabilisierende Wirkung haben. Die stabilisierende oder destabilisierende Wirkung muss von Fall zu Fall beurteilt werden, und es können nur wenige pauschale Aussagen gemacht werden.