
Aislamiento y caracterización de derivados de pterosina de P. aquilinum
Se aislaron derivados de pterosina de las plantas enteras de P. aquilinum. Para la investigación de los constituyentes fitoquímicos del helecho, se partió sucesivamente un extracto acuoso con acetato de etilo (EtOAc) y n-butanol (n-BuOH) (Fig. 1). La cromatografía en columna repetida de la fracción soluble en EtOAc dio como resultado el aislamiento de nueve derivados, incluyendo la (2R)-pterosina B (1), la pterosina Z (2), la (2S)-pterosina P (3) (3R)-pterosina D (4), (2S)-pterosina A (5), (2S,3R)-pterosina C (6), (2R,3R)-pterosina C (7), (2R)-pterosido B (8) y pterosido Z (9), con purezas superiores al 97% (Fig. 2). La repetida separación cromatográfica en columna de la fracción soluble en n-BuOH dio como resultado el aislamiento de seis derivados, incluyendo el (2S)-pterosido A (10), el (2S)-pterosido A2 (11), el (2S,3R)-pterosido C (12), el (2R,3R)-pterosido C (13), el (3S)-pterosido D (14), y el (2S)-pterosido P (15), con purezas superiores al 97%. Las estructuras de los compuestos se identificaron mediante el análisis de los datos espectrales, incluyendo MS, 1D- y 2D-NMR (Información suplementaria 1).

Estructuras de los compuestos de pterosina 1-15xxx
Actividad inhibitoria de los derivados de pterosina contra BACE1, AChE, y BChE
Para evaluar el potencial anti-AD, se evaluó la actividad inhibitoria de cada compuesto de pterosina frente a BACE1 y colinesterasas mediante los respectivos ensayos de inhibición in vitro (Tabla 1). Todos los derivados de pterosina probados mostraron actividades inhibitorias dependientes de la concentración contra BACE1 con un rango de valores IC50 (concentración inhibitoria media máxima) de 9,74-94,4 μM, con la excepción de la (2S)-pterosina A y el (2S)-pterosido P que fueron inactivos a las concentraciones probadas, en comparación con el IC50 de la quercetina utilizada como control positivo, que fue de 18,8 μM. La potencia inhibitoria de los inhibidores más potentes fue en el orden de (2R,3R)-pterosido C, (3S)-pterosido D, (2R)-pterosido B, (2S,3R)-pterosina C, (2R,3R)-pterosina C, (2S,3R)-pterosido C, y (2R)-pterosina B con valores de IC50 de 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 y 29,6 μM, respectivamente. Posteriormente, probamos los potenciales inhibitorios de los derivados de la pterosina contra la AChE. Todos los compuestos probados mostraron actividades inhibidoras de la AChE significativas, con valores de IC50 en el rango de 2,55-110 μM, en comparación con la IC50 contra la AChE de la berberina utilizada como control positivo, que fue de 0,39 μM. Los compuestos de pterosina que mostraron la mayor actividad inhibitoria contra la AChE fueron el (2R)-pterosido B, el (2R,3R)-pterosido C, el (2S,3R)-pterosido C, el (2S,3R)-pterosin C y el (2R)-pterosin B con valores de IC50 de 2,55, 3,77, 9,17, 12,8 y 16,2 μM, respectivamente. Por último, probamos las capacidades inhibitorias de los derivados de la pterosina contra la BChE. Todos los compuestos de pterosina ensayados mostraron actividad inhibitoria frente a la BChE, con valores de IC50 que oscilaron entre 5,29 y 119 μM, con la excepción de la (3R)-pterosina D que resultó inactiva a las concentraciones ensayadas, en comparación con la IC50 de la berberina frente a la BChE, que fue de 3,32 μM. Los compuestos de pterosina que mostraron la mayor actividad inhibitoria contra la BChE fueron el (2R,3R)-pterosido C y el pterosido Z con valores de IC50 de 5,29 y 5,31 μM, respectivamente.
En conjunto, la mayoría de los derivados de pterosina ensayados mostraron actividades inhibitorias significativas contra BACE1, AChE y BChE simultáneamente. La presencia del grupo adicional 2-hidroximetil-tetrahidro-pirano-3,4,5-triol como en los derivados de pterosida aumentó significativamente las actividades inhibitorias contra las enzimas. Además, la presencia del grupo hidroximetilo adicional en la posición 2 del anillo de indanona de la (2R)-pterosina B, como en la (2S)-pterosina A, o del grupo metilo, como en la (3R)-pterosina D y la pterosina Z, disminuyó las actividades inhibitorias contra las enzimas. En particular, la presencia del grupo hidroximetilo en la posición 5 del anillo de indanona, como en la (2S)-pterosina P, disminuyó la actividad inhibitoria frente a BACE1.
Parámetros cinéticos de la inhibición enzimática por parte de los derivados de la pterosina
En un intento de explicar el modo de inhibición enzimática de los derivados de la pterosina, realizamos un análisis cinético de BACE1 y las colinesterasas para los inhibidores representativos (Tabla 2, Información complementaria 2). Un Ki (constante de inhibición) bajo indica una unión más estrecha de la enzima y un inhibidor más eficaz. En general, los valores de Ki de los compuestos se correlacionaron con los respectivos valores de IC50. La inhibición de BACE1 por los compuestos (2R,3R)-pterosida C, (3S)-pterosida D y (2R,3R)-pterosina C fue de tipo mixto con valores de Ki de 12,6, 16,5 y 276 µM, respectivamente, mientras que la inhibición por el (2R)-pterosido B, la (2S,3R)-pterosina C y la (2R)-pterosina B fue no competitiva con valores de Ki de 23,1, 33,8 y 38,3 µM, respectivamente. La inhibición de la AChE por el (2R)-pterosido B, el (2R,3R)-pterosido C, la (2R)-pterosina B, la (2S,3R)-pterosina C y el (3S)-pterosido D fue de tipo mixto con valores de Ki de 489, 8,13, 12,1, 16,3 y 23,1 µM, respectivamente, mientras que la (2R,3R)-pterosina C fue un inhibidor de tipo no competitivo con un valor Ki de 29,6 µM. La inhibición de la BChE por la (2R,3R)-pterosina C, el (2R,3R)-pterosido C, el (3S)-pterosido D y el (2R)-pterosido B fue de tipo mixto con valores de Ki de 4,77, 9,62, 19.7 y 22,6 µM, respectivamente, mientras que la (2S,3R)-pterosina C y la (2R)-pterosina B fueron inhibidores no competitivos con valores de Ki de 29,9 y 53,5 µM, respectivamente. Así pues, estos resultados sugieren que los derivados específicos de la pterosina podrían ser inhibidores eficaces de BACE1, AChE y BChE.
Simulaciones de acoplamiento molecular para BACE1, AChE y BChE
Se dispone de varias estructuras cristalinas para BACE1 y colinesterasas. Seleccionamos los PDB humanos basándonos en las estructuras de tipo salvaje, los ligandos cocristalizados y las resoluciones de las estructuras. Se seleccionaron para el acoplamiento las estructuras de cristal de rayos X de BACE1 en complejo con QUD (código PDB: 2WJO, resolución: 2,5 Å)33 , de AChE en complejo con E2020 (código PDB: 4EY7, resolución: 2,35 Å)34 , y de BChE en complejo con 3F9 (código PDB: 4TPK, resolución: 2,70 Å)35 . Inicialmente, QUD, E2020 y 3F9 fueron extraídos de las estructuras cristalinas y redocked en los sitios activos de BACE1, AChE y BChE, respectivamente. Posteriormente, se acoplaron el (2R,3R)-pterosido C, el (3S)-pterosido D, el (2R,3R)-pterosin C, el (2R)-pterosido B, el (2S,3R)-pterosin C y el (2R)-pterosin B con el mecanismo conocido de inhibición contra BACE1, AChE y BChE para determinar su SAR. Los resultados del docking se resumen en la Tabla 3. El SAR de los inhibidores seleccionados de tipo mixto y no competitivo de BACE1, AChE y BChE permitió evaluar la nueva inhibición dirigida al sitio de unión de las enzimas por las pterosinas.
Nuestro modo de acoplamiento de E2020 fue consistente con el modo de unión determinado experimentalmente reportado previamente con AChE humana recombinante (rhAChE) (Información Suplementaria 3)34. La desviación media cuadrática (RMSD) entre el cristal y las conformaciones acopladas de E2020 fue de 0,54 Å, lo que sugiere la fiabilidad de nuestra configuración de acoplamiento para reproducir el modo de unión experimental. Además, el modo acoplado de E2020 condujo a una interacción similar a la de rhAChE-E2020. En nuestro estudio, las moléculas de agua se eliminaron de la estructura cristalina durante el acoplamiento; por lo tanto, las interacciones mediadas por el agua no se analizaron en el presente estudio. Del mismo modo, los modos de acoplamiento de QUD y 3F9 eran consistentes con los datos experimentales disponibles para BACE1 33 y BChE35, respectivamente (Información suplementaria 3). Los RMSDs entre el cristal y las conformaciones acopladas de QUD y 3F9 fueron de 0,46 y 0,60 Å, respectivamente. Además, los sitios de unión de los inhibidores de la pterosina estaban de acuerdo con un estudio de acoplamiento previo que incluía a BACE1, AChE y BChE38. Sin embargo, el estudio utilizó la AChE de Tetronarce californica (código PDB: 1ACJ), que contiene números de residuos ligeramente diferentes a los de la AChE humana debido a las variaciones en sus secuencias.
Acoplamiento de BACE1
Basado en el tipo de inhibición y la actividad, se seleccionaron el (2R,3R)-pterosido C y el (2R)-pterosido B para demostrar los modos acoplados de los inhibidores de tipo mixto y no competitivo de BACE1, respectivamente. La Figura 3a, b muestra los modelos de acoplamiento del (2R,3R)-pterosido C y del (2R)-pterosido B, respectivamente. Las interacciones de los compuestos acoplados dentro del sitio activo de BACE1 se muestran en la Fig. 4.

Molecular docking models for a the mixed-type BACE1 inhibitor (2R,3R)-pteroside C (blue color) and b the noncompetitive BACE1 inhibitor (2R)-pteroside B (brown color). Las poses acopladas se superponen a la estructura cristalina de rayos X de QUD (color amarillo) (código PDB: 2WJO). BACE1, los residuos del sitio activo y los compuestos se muestran mediante modelos de cinta, línea y palo, respectivamente. Los colores de las líneas punteadas explican los tipos de diversas interacciones: interacciones de enlace de hidrógeno (verde), interacciones hidrofóbicas (rosa) e interacciones π-sigma (púrpura). BACE1 β-sitio de la proteína precursora de amiloide que escinde la enzima 1

Diagrama de interacción de ligandos de a QUD, b (2R,3R)-pterosida C, c (3S)-pterosida D, d (2R,3R)-pterosina C, e (2R)-pterosida B, f (2S,3R)-pterosina C, y g (2R)-pterosina B en el sitio activo de BACE1. Los colores de las líneas punteadas explican los tipos de diversas interacciones: interacciones de enlace de hidrógeno (verde), interacciones hidrofóbicas (rosa) e interacciones π-sigma (púrpura). BACE1 β-site amyloid precursor protein cleaving enzyme 1
La pose acoplada de QUD mostró una energía de unión (B.E.) de -7,59 kcal/mol. Como se muestra en la Fig. 4a, el grupo NH2 del anillo de quinazolina del ligando mostró dos enlaces de hidrógeno con los grupos CO de Asp32 y Asp228 a distancias de 1,86 y 2,17 Å, respectivamente. Se observó un tercer enlace de hidrógeno entre el otro grupo NH del ligando y el grupo CO de Gly230 a una distancia de 2,16 Å. Leu30, Tyr71, Phe108 y Val332 mediaron las interacciones hidrofóbicas. La figura 4b-d muestra las posturas acopladas del (2R,3R)-pterosido C, (3S)-pterosido D y (2R,3R)-pterosina C (inhibidores de tipo mixto de BACE1), respectivamente. Se posicionaron en el bolsillo de unión revestido por Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 y Thr231. En cuanto a sus niveles de actividad, el (2R,3R)-pterosido C (IC50 = 9,74 µM), el (3S)-pterosido D (IC50 = 10,7 µM) y la (2R,3R)-pterosina C (IC50 = 26,2 µM) mostraron un E.B. de -7,27, -6,93 y -4,84 kcal/mol, respectivamente. El (2R,3R)-pterosido C mostró una mayor potencia que la (2R,3R)-pterosina C debido a la existencia de un grupo adicional 2-hidroximetil-tetrahidro-pirano-3,4,5-triol, que formó tres enlaces de hidrógeno (Fig. 4b). El grupo OH del hidroximetilo y el grupo 3-OH del anillo tetrahidro-pirano-triol mostraron dos enlaces de hidrógeno con el grupo CO de Asp228 a distancias de 2,63 y 2,12 Å, respectivamente. Además, el grupo 3-OH mostró un enlace de hidrógeno con Thr231 a una distancia de 1,89 Å. En el caso del (3S)-pterosido D (Fig. 4c), la presencia del grupo 2,2-dimetil en el anillo de indanona alteró ligeramente las interacciones de unión en comparación con el (2R,3R)-pterosido C. El grupo 3-OH del anillo tetrahidro-pirano-triol mostró un único enlace de hidrógeno con Asp228 a una distancia de 1,93 Å. Sin embargo, el grupo 3-OH del anillo de indanona mostró un enlace de hidrógeno adicional con Ile126 a una distancia de 2,34 Å. Estas interacciones redujeron ligeramente la actividad del (3S)-pterosido D en comparación con el (2R,3R)-pterosido C. Las interacciones de acoplamiento de la (2R,3R)-pterosina C mostradas en la Fig. 4d muestran el grupo 3-OH del anillo de indanona unido a Ser36 y Asn37 mediante dos enlaces de hidrógeno a distancias de 1,83 y 2,07 Å, respectivamente. Ala39, Val69, Trp76, Ile118 y Arg128 participaron en interacciones hidrofóbicas, mientras que Val69 mostró una interacción π-sigma.
Las poses acopladas de (2R)-pterosida B, (2S,3R)-pterosina C y (2R)-pterosina B (inhibidores no competitivos de BACE1) se muestran en la Fig. 4e-g, respectivamente. Se acoplaron en la cavidad delimitada por Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 e Ile126. De acuerdo con sus valores de actividad, la (2R)-pterosina B (IC50 = 18,0 µM), la (2S,3R)-pterosina C (IC50 = 23,1 µM) y la (2R)-pterosina B (IC50 = 29,6 µM) mostraron una E.B. de -6,16, -5,07 y -4,64 kcal/mol, respectivamente. Como se muestra en la Fig. 4e, el (2R)-pteroside B demostró una mayor actividad que el (2S,3R)-pterosin C y el (2R)-pterosin B debido a la presencia de un grupo adicional de 2-hidroximetil-tetrahidro-pirano-3,4,5-triol, que mostró cuatro interacciones de enlace de hidrógeno. Se observaron dos enlaces de hidrógeno entre el grupo 4-OH del anillo tetrahidro-pirano-triol y los grupos NH y CO de la Asn37 a distancias de 2,48 y 2,19 Å, respectivamente. Además, los grupos 3 y 5-OH mostraron dos enlaces de hidrógeno adicionales con Ile126 y Trp76 a distancias de 2,19 y 2,46 Å, respectivamente. La (2S,3R)-pterosina C mostró una actividad ligeramente mejor que la (2R)-pterosina B debido a la presencia de un grupo OH adicional en la posición 3 del anillo de indanona, que formó un enlace de hidrógeno con Lys107 a una distancia de 2,12 Å (Fig. 4f). Las demás interacciones fueron similares a las de la (2R)-pterosina B. Como se muestra en la Fig. 4g, la (2R)-pterosina B mostró interacciones hidrofóbicas con Val69, Tyr76 y Phe108.
Docking de AChE
(2R)-Pterosida B y (2R,3R)-pterosina C fueron seleccionados como representantes para demostrar los modos de acoplamiento de los inhibidores de AChE de tipo mixto y no competitivo, respectivamente, debido a sus actividades y tipo de inhibición de AChE. La Figura 5a, b ilustra los modelos de acoplamiento del (2R)-pterosido B y del (2R,3R)-pterosin C, respectivamente. Las interacciones de los compuestos acoplados dentro del sitio activo de la AChE se muestran en la Fig. 6.

Modelos de acoplamiento molecular para a el inhibidor de la AChE de tipo mixto (2R)-pterosina B (color azul) y b el inhibidor no competitivo de la AChE (2R,3R)-pterosina C (color marrón). Las posturas acopladas se superponen a la estructura cristalina de rayos X de E2020 (color amarillo) (código PDB: 4EY7). La AChE, los residuos del sitio activo y los compuestos se muestran mediante modelos de cinta, línea y palo, respectivamente. Los colores de las líneas punteadas explican los tipos de diversas interacciones: interacciones de enlace de hidrógeno (verde) e interacciones hidrofóbicas (rosa). AChE acetilcolinesterasa

Diagrama de interacción de ligandos de a E2020, b (2R)-pterosida B, c (2R,3R)-pterosida C, d (2S,3R)-pterosina C, e (2R)-pterosina B, f (3S)-pterosida D y g (2R,3R)-pterosina C en el sitio activo de AChE. Los colores de las líneas punteadas explican los tipos de diversas interacciones: interacciones de enlace de hidrógeno (verde), interacciones hidrofóbicas (rosa) e interacciones π-sigma (púrpura). AChE acetilcolinesterasa
La pose acoplada de E2020 demostró un B.E. de -10,28 kcal/mol. Como se ilustra en la Fig. 6a, el grupo CO del anillo de indanona formó un enlace de hidrógeno con el grupo NH de Phe295 a una distancia de 1,70 Å. Trp286 y Tyr341 participaron en interacciones π-sigma, mientras que Trp86, Trp286, Tyr337, Phe338 y Tyr341 mediaron interacciones hidrofóbicas. La figura 6b-f muestra las poses acopladas del (2R)-pterosido B, (2R,3R)-pterosido C, (2S,3R)-pterosina C, (2R)-pterosina B y (3S)-pterosido D (inhibidores de la AChE de tipo mixto), respectivamente. Se alojaron en el sitio activo rodeados de Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 y Tyr341. En consonancia con sus valores de actividad, el (2R)-pterosido B (IC50 = 2,55 µM), el (2R,3R)-pterosido C (IC50 = 3,77 µM), la (2S,3R)-pterosina C (IC50 = 12.8 µM), la (2R)-pterosina B (IC50 = 16,2 µM) y el (3S)-pterosido D (IC50 = 27,4 µM) mostraron una E.B. de -7,90, -7,49, -6,03, -5,76 y -4,91 kcal/mol, respectivamente. El (2R)-pterosido B demostró una mayor potencia que la (2S,3R)-pterosina C y la (2R)-pterosina B debido a la presencia de un grupo adicional 2-hidroximetil-tetrahidro-pirano-3,4,5-triol, que estableció tres interacciones de enlace de hidrógeno (Fig. 6b). El grupo OH del hidroximetilo y el grupo 3-OH del anillo tetrahidro-pirano-triol mostraron enlaces de hidrógeno con el grupo CO de Asn87 a distancias de 2,23 y 2,28 Å, respectivamente. Además, el grupo 4-OH formó un enlace de hidrógeno con el grupo CO de Trp86 a una distancia de 2,13 Å. En el caso del (2R,3R)-pterosido C (Fig. 6c), el grupo metilo en la posición 2 del anillo de indanona no mostró una interacción hidrofóbica con Tyr341 y, por tanto, mostró una actividad comparativamente menor que el (2R)-pterosido B. Sin embargo, Tyr341 mantuvo la interacción hidrofóbica con la otra parte del anillo de indanona, como se muestra en la (2R)-pterosina B. La mayor actividad de la (2S,3R)-pterosina C que la (2R)-pterosina B se atribuyó a la existencia de un grupo OH adicional en la posición-3 del anillo de indanona, que formó un enlace de hidrógeno con Phe295 a una distancia de 1,82 Å (Fig. 6d). El resto de interacciones fueron comparables a las de la (2R)-pterosina B. Como se muestra en la Fig. 6e, el grupo OH del grupo hidroxietil en la posición 6 del anillo de indanona formó un enlace de hidrógeno con Tyr124 a una distancia de 2,48 Å. Tyr124, Trp286, Tyr337, Phe338 y Tyr341 contribuyeron a las interacciones hidrofóbicas. En comparación con el (2R)-pterosido B y el (2R,3R)-pterosido C, el (3S)-pterosido D mostró interacciones de unión diferentes debido a la presencia del grupo 2,2-dimetil en el anillo de indanona (Fig. 6f). El grupo 2,2-dimetil contribuyó de forma significativa a la distinta postura de acoplamiento del (3S)-pterosido D. El grupo 5-OH del anillo tetrahidro-pirano-triol formó un enlace de hidrógeno con Asp74 a una distancia de 1,96 Å. El grupo CO del anillo de indanona mostró un enlace de hidrógeno con Tyr72 a una distancia de 2,97 Å. Estas interacciones explican la baja actividad del (3S)-pterosido D.
La pose acoplada de la (2R,3R)-pterosina C (inhibidor no competitivo de la AChE) se muestra en la Fig. 6g. La (2R,3R)-pterosina C (IC50 = 23,2 µM) demostró un B.E. de -5,01 kcal/mol. El bolsillo de unión de la (2R,3R)-pterosina C comprendía Tyr72, Trp286, Ser293, Phe297 y Tyr341, con dos interacciones de enlace de hidrógeno. Uno de los enlaces de hidrógeno se formó entre el grupo 3-OH del anillo de indanona y el grupo CO de Ser293 a una distancia de 2,03 Å. El segundo enlace de hidrógeno se observó entre el grupo OH del grupo hidroxietil presente en la posición 6 del anillo de indanona y Tyr72 a una distancia de 1,94 Å. Residuos como Tyr72, Trp286, Phe297 y Tyr341 participaron en las interacciones hidrofóbicas.
Docking de la BChE
Considerando los niveles de actividad y el tipo de inhibición de la BChE, se seleccionaron el (2R,3R)-pterosido C y la (2S,3R)-pterosina C para demostrar los modos acoplados de los inhibidores de la BChE de tipo mixto y no competitivo, respectivamente. La figura 7a, b ilustra los modelos de acoplamiento del (2R,3R)-pterosido C y del (2S,3R)-pterosin C, respectivamente. Las interacciones de los compuestos acoplados dentro del sitio activo de la BChE se presentan en la Fig. 8.
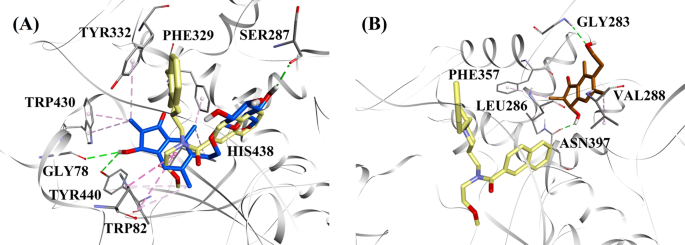
Modelos de acoplamiento molecular para a el inhibidor de BChE de tipo mixto (2R,3R)-pterosina C (color azul) y b el inhibidor de BChE no competitivo (2S,3R)-pterosina C (color marrón). Las posturas acopladas se superponen a la estructura cristalina de rayos X de 3F9 (color amarillo) (código PDB: 4TPK). La BChE, los residuos del sitio activo y los compuestos se muestran mediante modelos de cinta, línea y palo, respectivamente. Los colores de las líneas de puntos explican los tipos de diversas interacciones: interacciones de enlace de hidrógeno (verde) e interacciones hidrofóbicas (rosa). BChE butirilcolinesterasa
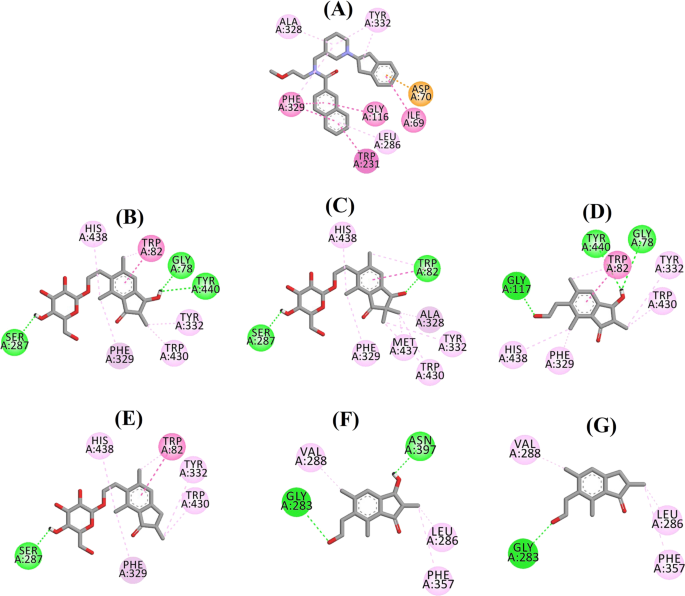
Diagrama de interacción de ligandos de a 3F9, b (2R,3R)-pterosida C, c (3S)-pterosida D, d (2R,3R)-pterosina C, e (2R)-pterosida B, f (2S,3R)-pterosina C y g (2R)-pterosina B en el sitio activo de BChE. Los colores de las líneas punteadas explican los tipos de diversas interacciones: interacciones de enlace de hidrógeno (verde), interacciones hidrofóbicas (rosa) e interacciones π-aniónicas (dorado). BChE butirilcolinesterasa
La pose acoplada de 3F9 mostró un B.E. de -8,49 kcal/mol. Como se muestra en la Fig. 8a, las interacciones hidrofóbicas fueron las principales responsables de la unión del ligando. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 y Tyr332 fueron responsables de las interacciones hidrofóbicas, mientras que Asp70 demostró una interacción π-aniónica. La figura 8b-e ilustra las poses acopladas del (2R,3R)-pterosido C, el (3S)-pterosido D, el (2R,3R)-pterosin C y el (2R)-pterosido B (inhibidores de la BChE de tipo mixto), respectivamente. Su bolsillo de unión estaba compuesto por Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 y Tyr440. De acuerdo con sus niveles de actividad, el (2R,3R)-pterósido C (IC50 = 5,29 µM), el (3S)-pterósido D (IC50 = 19,3 µM), el (2R,3R)-pterósido C (IC50 = 20,3 µM) y el (2R)-pterósido B (IC50 = 62,0 µM) demostraron una E.B. de -7,23, -6,59, -6,52 y -4,38 kcal/mol, respectivamente. Como se muestra en la Fig. 8b, el grupo 3-OH del anillo de indanona del (2R,3R)-pterosido C mostró dos enlaces de hidrógeno con Gly78 y Tyr440 a distancias de 2,87 y 2,89 Å, respectivamente. En el caso del (3S)-pterosido D, el grupo 2,2-dimetil en el anillo de indanona afectó a las interacciones de unión del grupo 3-OH (Fig. 8c). El grupo 3-OH sólo formó un enlace de hidrógeno con Trp82 a una distancia de 2,94 Å, lo que dio lugar a una actividad comparativamente menor del (3S)-pterosida D que del (2R,3R)-pterosida C. Como se muestra en la Fig. 8d, la (2R,3R)-pterosina C no pudo producir un enlace de hidrógeno con Ser287 debido a la ausencia de un grupo 2-hidroximetil-tetrahidro-pirano-3,4,5-triol. En consecuencia, su actividad fue menor que la del (2R,3R)-pterosido C. La falta del grupo OH en la posición 3 del anillo de indanona fue la responsable de la escasa actividad del (2R)-pterosido B (Fig. 8e), que no pudo formar enlaces de hidrógeno con Gly78 y Tyr440 de forma similar al (2R,3R)-pterosido C.
Las poses acopladas del (2S,3R)-pterosin C y (2R)-pterosin B (inhibidores no competitivos de BChE) se muestran en la Fig. 8f, g, respectivamente. Estas poses acopladas estaban contenidas en la cavidad encerrada por Gly283, Leu286, Val288, Phe357 y Asn397. Según sus niveles de actividad, la (2S,3R)-pterosina C (IC50 = 44,3 µM) y la (2R)-pterosina B (IC50 = 48,1 µM) mostraron un E.B. de -5,40 y -5,06 kcal/mol, respectivamente. La (2S,3 R)-pterosina C fue más potente que la (2R)-pterosina B debido a la presencia de un grupo OH adicional en la posición 3 del anillo de indanona, que formó un enlace de hidrógeno con el grupo CO de la Asn397 a una distancia de 2,05 Å (Fig. 8f). Otras interacciones fueron similares a la (2R)-pterosina B. Como se muestra en la Fig. 8g, el grupo OH del grupo hidroxietilo en la posición 6 del anillo de indanona formó un enlace de hidrógeno con Gly283 a una distancia de 2,40 Å. Los residuos Leu286, Val288 y Phe357 participaron en interacciones hidrofóbicas.
Los inhibidores de tipo mixto se unen tanto a la enzima libre como al complejo enzima-sustrato, lo que indica que estos compuestos pueden unirse al sitio catalítico de cada enzima correspondiente. Los inhibidores no competitivos se unen al sitio alostérico de la enzima libre o del complejo enzima-sustrato. Un estudio reciente sugirió que los inhibidores competitivos, de tipo mixto y no competitivos ocupan diferentes sitios en los bolsillos de unión de BACE1, AChE y BChE38. Durante el docking para la evaluación del mecanismo inhibitorio de los derivados de la pterosina, se definieron los sitios de unión de los compuestos según su tipo de inhibición. Los resultados del acoplamiento indicaron que los sitios de unión de los inhibidores de tipo mixto y no competitivo para BACE1, AChE y BChE se solapan parcialmente en cada sitio activo correspondiente y son coherentes con un informe anterior38.
Permeabilidad de la BBB
PAMPA-BBB, un ensayo de permeabilidad de la membrana artificial in vitro para la BBB, es una de las herramientas de cribado fisicoquímico más fiables en la fase inicial de descubrimiento de fármacos dirigidos al SNC40. El sistema PAMPA-BBB modela la difusión pasiva transcelular de sustancias químicas a través de la BBB y mide los mecanismos de transporte estrictamente pasivos a través de una membrana lipídica artificial en la permeabilidad efectiva (Pe, cm/s). Sobre la base del patrón establecido para la predicción de la permeabilidad BBB, los compuestos se clasificaron en (i) «CNS+» (alta permeabilidad BBB prevista); Pe (10-6 cm/s) > 4,00, (ii) «CNS-» (baja permeabilidad BBB prevista); Pe (10-6 cm/s) < 2,00, y (iii) «CNS+/-» (permeabilidad BBB incierta); Pe (10-6 cm/s) de 4,00 a 2,00. En consecuencia, la (2R)-pterosina B, la (2S)-pterosina P y la (2S)-pterosina A mostraron una alta permeabilidad BBB con valores de Pe de 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s y 6,26 × 10-6 cm/s, respectivamente (Tabla 4, Información complementaria 4). El valor Pe de la (2R)-pterosina B fue 1,7 veces mayor que el del fármaco del SNC verapamilo (Pe = 34,6 × 10-6 cm/s), que se utilizó como control positivo. La (2S,3R)-pterosina C y la (2R,3R)-pterosina C mostraron una permeabilidad BBB incierta con valores de Pe de 2,34 y 1,98, respectivamente. El (2R,3R)-pterosido C, el (3S)-pterosido D y el (2R)-pterosido B, que mostraron las actividades inhibidoras de la BACE1 y la colinesterasa más potentes entre los derivados de la pterosina probados, mostraron una permeabilidad BBB muy baja. La existencia del grupo 2-hidroximetil-tetrahidro-pirano-3,4,5-triol, como en los derivados de pterosina, dio lugar a una notable disminución de la permeabilidad de la BBB. En comparación con la (2R)-pterosina B, la presencia adicional del grupo OH en la posición 3 del anillo de indanona como en la pterosina C, el grupo hidroximetil en la posición 2 del anillo de indanona como en la (2S)-pterosina A o el grupo hidroximetil en la posición 5 del anillo de indanona como en la (2S)-pterosina P también redujo significativamente la permeabilidad de la BBB. Teniendo en cuenta una permeabilidad de la BBB excepcionalmente alta y la inhibición significativa de BACE1, AChE y BChE, la (2R)-pterosina B puede tener el potencial de exhibir una fuerte actividad anti-AD.
Efectos de la (2R)-pterosina B y del (2R,3R)-pterosido C sobre la secreción de péptidos Aβ por parte de las células neuronales
Para investigar la función de la (2R)-pterosina B y del (2R,3R)-pterosida C en la disminución de la excreción de Aβ de las células neuronales, utilizamos una línea celular de neuroblastoma murino que sobreexpresa de forma estable la APPswe humana. La línea celular es un modelo celular de EA caracterizado por la secreción excesiva de Aβ40 y Aβ42. Los oligómeros amiloides tóxicos se forman a partir de las dos isoformas del péptido Aβ con diferentes longitudes. Aβ40 es la isoforma Aβ más abundante en el cerebro41, mientras que Aβ42 aumenta significativamente con ciertas formas de EA42. El ELISA de sándwich de Aβ40 mostró que la (2R)-pterosina B redujo significativamente la cantidad de péptido Aβ40 secretado por las células de neuroblastoma en el medio hasta un 50% a 500 μM (P < 0,01) (Fig. 9a). Del mismo modo, la secreción del péptido Aβ42 por parte de las células de neuroblastoma disminuyó significativamente en presencia de 500 µM de (2R,3R)-pterosida C (P < 0,05) (Fig. 9b). En conclusión, la (2R)-pterosina B y el (2R,3R)-pterosido C disminuyeron significativamente la secreción de péptidos Aβ de las células de neuroblastoma a una concentración de 500 μM.

a Efecto de (2R)-pterosina B sobre la secreción de péptidos Aβ. Las células de neuroblastoma secretoras de APPswe fueron tratadas con 12, 60, 250 o 500 µM de (2R)-pterosina B durante 24 h, y se recogieron los medios condicionados en presencia de un inhibidor de la proteasa. Se utilizó β-SI (10 µM) como control positivo. El control negativo incluyó células cultivadas en ausencia de compuestos de prueba. El análisis cuantitativo de Aβ40 y Aβ42 secretados en los medios condicionados se realizó mediante ELISA tipo sándwich. Se muestran las medias ± SD de tres experimentos independientes. Los péptidos Aβ40 secretados disminuyeron significativamente en presencia de 500 µM de (2R)-pterosina B. Control negativo CON, β-SI β-inhibidor de la secretasa III, Aβ β-amiloide. *P < 0,05, **P < 0,01. b Efecto de la (2R,3R)-pterosina C sobre la secreción de péptidos Aβ. Las células de neuroblastoma secretoras de APPswe fueron tratadas con 12, 60, 125, 250 o 500 µM de (2R,3R)-pterosida C durante 24 h, y se recogieron los medios condicionados en presencia de un inhibidor de la proteasa. Se utilizó β-SI (10 µM) como control positivo. El control negativo incluyó células cultivadas en ausencia de los compuestos de prueba. El análisis cuantitativo de Aβ40 y Aβ42 secretados en los medios condicionados se realizó mediante ELISA de sándwich. Se muestran las medias ± SD de tres experimentos independientes. El péptido Aβ42 secretado disminuyó significativamente en presencia de 500 µM de (2R,3R)-pterosida C. Control negativo CON, β-SI β-inhibidor de la secretasa III, Aβ β-amiloide. *P < 0.05
Citotoxicidad de los derivados de la pterosina basada en el ensayo MTT43
En general, los derivados de la pterosina mostraron una citotoxicidad insignificante contra varias líneas celulares normales y cancerosas, como el neuroblastoma humano SH-SY5Y, las células gliales de rata C6, los fibroblastos embrionarios de ratón NIH3T3 y el melanoma de ratón B16F10 con valores de LD50 superiores a 0.5 mM (Información complementaria 5). En particular, la (2R)-pterosina B, la (3R)-pterosina D, la (2S)-pterosina P, el (2S)-pterosido A, el (2R)-pterosido B y el (2R,3R)-pterosido C no mostraron citotoxicidad contra las líneas celulares analizadas con valores de LD50 superiores a 5 mM. Curiosamente, varias pterosinas mostraron efectos antiproliferativos relativos contra las células neuronales SH-SY5Y en comparación con las células gliales C6 de origen mesenquimal. Los presentes resultados concuerdan con un informe anterior que sugiere que los derivados de la pterosina no son tóxicos para los seres humanos26.