
Isolation and characterization of pterosin derivatives from P. aquilinum
Pterozin-származékokat izoláltunk a P. aquilinum egész növényéből. A páfrányfenyőből származó fitokémiai összetevők vizsgálatához a vizes kivonatot egymás után etil-acetáttal (EtOAc) és n-butanollal (n-BuOH) osztottuk (1. ábra). Az EtOAc-ban oldódó frakció ismételt oszlopkromatográfiája kilenc származék izolálását eredményezte, köztük a (2R)-pterozin B (1), pterozin Z (2), (2S)-pterozin P (3), (3R)-pterozin D (4), (2S)-pterozin A (5), (2S,3R)-pterozin C (6), (2R,3R)-pterozin C (7), (2R)-pterozid B (8) és pterozid Z (9), 97%-nál nagyobb tisztasággal (ábra). 2). Az n-BuOH-ban oldódó frakció ismételt oszlopkromatográfiás elválasztása hat származék, köztük a (2S)-pterozid A (10), (2S)-pterozid A2 (11), (2S,3R)-pterozid C (12), (2R,3R)-pterozid C (13), (3S)-pterozid D (14) és (2S)-pterozid P (15) izolálását eredményezte 97%-nál nagyobb tisztasággal. A vegyületek szerkezetét a spektrális adatok elemzésével azonosítottuk, beleértve az MS, 1D- és 2D-NMR adatokat (1. kiegészítő információ).

A pterozin vegyületek 1-15xxx szerkezetei
A pterozin származékok BACE1, AChE elleni gátló hatása, és BChE
Az AD-ellenes potenciál értékeléséhez az egyes pterozinvegyületek BACE1 és kolinészterázok elleni gátló hatását a megfelelő in vitro gátlási vizsgálatokkal értékeltük (1. táblázat). Minden vizsgált pterozin-származék koncentrációfüggő gátló hatást mutatott a BACE1 ellen, az IC50 értékek (félmaximális gátló koncentráció) tartománya 9,74-94,4 μM volt, a (2S)-pterozin A és a (2S)-pterozid P kivételével, amelyek a vizsgált koncentrációkban inaktívak voltak, szemben a pozitív kontrollként használt kvercetin IC50 értékével, amely 18,8 μM volt. A legerősebb inhibitorok gátló ereje a következő sorrendben volt: (2R,3R)-pterozid C, (3S)-pterozid D, (2R)-pterozid B, (2S,3R)-pterozin C, (2R,3R)-pterozin C, (2S,3R)-pterozid C és (2R)-pterozin B, 9 IC50 értékkel.74, 10,7, 18,0, 23,1, 26,2, 28,9 és 29,6 μM. Ezt követően megvizsgáltuk a pterozin-származékok AChE elleni gátló potenciálját. Valamennyi vizsgált vegyület jelentős AChE-gátló aktivitást mutatott, az IC50 értékek 2,55-110 μM tartományban voltak, szemben a pozitív kontrollként használt berberin AChE elleni IC50 értékével, amely 0,39 μM volt. Az AChE-vel szemben a legerősebb gátló hatást mutató pterozin vegyületek a (2R)-pterozid B, (2R,3R)-pterozid C, (2S,3R)-pterozid C, (2S,3R)-pterozin C és (2R)-pterozin B voltak, 2,55, 3,77, 9,17, 12,8 és 16,2 μM IC50 értékekkel. Végül a pterozin-származékok BChE elleni gátló képességét vizsgáltuk. Minden vizsgált pterozin vegyület gátló hatást mutatott a BChE-vel szemben, az IC50 értékek 5,29 és 119 μM között mozogtak, kivéve a (3R)-pterozin D-t, amely a vizsgált koncentrációkban inaktív volt, szemben a berberin BChE-vel szembeni IC50 értékével, amely 3,32 μM volt. A BChE-vel szemben a legerősebb gátló hatást mutató pterozin vegyületek a (2R,3R)-pterozid C és a pterozid Z voltak 5,29, illetve 5,31 μM IC50 értékkel.
A legtöbb vizsgált pterozin-származék együttesen jelentős gátló hatást mutatott a BACE1, AChE és BChE ellen egyszerre. A pterozid-származékokhoz hasonlóan a további 2-hidroximetil-tetrahidro-pirán-3,4,5-triol csoport jelenléte jelentősen növelte az enzimekkel szembeni gátló aktivitást. Ezenkívül a (2R)-pterozin B indanongyűrűjének 2. pozíciójában lévő további hidroximetilcsoport jelenléte, mint a (2S)-pterozin A-ban, vagy a metilcsoport jelenléte, mint a (3R)-pterozin D-ben és a pterozin Z-ben, csökkentette az enzimekkel szembeni gátló aktivitást. Különösen a hidroxi-metilcsoport jelenléte az indanongyűrű 5. pozíciójában, mint a (2S)-pterozin P-ben, csökkentette a BACE1 elleni gátló aktivitást.
A pterozin-származékok általi enzimgátlás kinetikai paraméterei
A pterozin-származékok enzimgátlási módjának magyarázatára tett kísérletként elvégeztük a BACE1 és a kolinészterázok kinetikai elemzését reprezentatív inhibitorokra (2. táblázat, 2. kiegészítő információ). Az alacsony Ki (gátlási állandó) szorosabb enzimkötésre és hatékonyabb inhibitorra utal. Összességében a vegyületek Ki értékei korreláltak a megfelelő IC50 értékekkel. A (2R,3R)-terozid C, a (3S)-terozid D és a (2R,3R)-terozin C vegyületek által okozott BACE1-gátlás vegyes típusú volt, 12,6, 16,5 és 27 Ki értékekkel.6 µM, míg a (2R)-pterozid B, (2S,3R)-pterozin C és (2R)-pterozin B általi gátlás nem kompetitív volt 23,1, 33,8 és 38,3 µM Ki értékekkel. Az AChE gátlása a (2R)-pterozid B, (2R,3R)-pterozid C, (2R)-pterozin B, (2S,3R)-pterozin C és (3S)-pterozid D által vegyes típusú volt 4 Ki értékkel.89, 8,13, 12,1, 16,3 és 23,1 µM, míg a (2R,3R)-pterozin C nem kompetitív típusú inhibitor volt 29,6 µM Ki értékkel. A BChE gátlása a (2R,3R)-pterozin C, a (2R,3R)-pterozid C, a (3S)-pterozid D és a (2R)-pterozid B által vegyes típusú volt, 4,77, 9,62, 19 Ki értékekkel.7, illetve 22,6 µM, míg a (2S,3R)-pterozin C és a (2R)-pterozin B nem kompetitív inhibitorok voltak 29,9, illetve 53,5 µM Ki értékkel. Így ezek az eredmények azt sugallták, hogy a specifikus pterozin-származékok hatékony BACE1, AChE és BChE gátlók lehetnek.
Molekuláris dokkolási szimulációk a BACE1, AChE és BChE esetében
A BACE1 és a kolinészterázok esetében több kristályszerkezet is rendelkezésre áll. A humán PDB-ket a vad típusú szerkezetek, az együtt kristályosított ligandumok és a szerkezetek felbontása alapján választottuk ki. A dokkoláshoz a QUD-dal komplexált BACE1 (PDB-kód: 2WJO, felbontás: 2,5 Å)33, az E2020-val komplexált AChE (PDB-kód: 4EY7, felbontás: 2,35 Å)34 és a 3F9-cel komplexált BChE (PDB-kód: 4TPK, felbontás: 2,70 Å)35 röntgenkristályszerkezeteit választottuk ki. Kezdetben a QUD-ot, az E2020-t és a 3F9-et kivontuk a kristályszerkezetekből, és redokkoltuk a BACE1, az AChE és a BChE aktív helyére. Ezt követően a (2R,3R)-pterozid C, (3S)-pterozid D, (2R,3R)-pterozin C, (2R)-pterozid B, (2S,3R)-pterozin C és (2R)-pterozin B ismert gátlási mechanizmusával a BACE1, AChE és BChE ellen dokkoltuk a SAR meghatározásához. A dokkolási eredményeket a 3. táblázat foglalja össze. A kiválasztott vegyes típusú és nem kompetitív BACE1, AChE és BChE inhibitorok SAR-ja lehetővé tette az enzimek pterozinok általi új kötőhely-irányított gátlásának értékelését.
Az E2020 dokkolási módja összhangban volt a rekombináns humán AChE (rhAChE) esetében korábban bejelentett, kísérletileg meghatározott kötési móddal (Kiegészítő információk 3)34. Az E2020 kristályos és dokkolt konformációja közötti átlagos négyzetes eltérés (RMSD) 0,54 Å volt, ami azt sugallta, hogy a dokkolási beállításunk megbízhatóan reprodukálja a kísérleti kötődési módot. Ezenkívül az E2020 dokkolt módja hasonló kölcsönhatáshoz vezetett, mint a rhAChE-E2020-é. Vizsgálatunkban a dokkolás során a vízmolekulákat eltávolítottuk a kristályszerkezetből, ezért a víz által közvetített kölcsönhatásokat jelen tanulmányban nem elemeztük. Hasonlóképpen, a QUD és a 3F9 dokkolt móduszai összhangban voltak a BACE1 33 és a BChE35 esetében rendelkezésre álló kísérleti adatokkal (3. kiegészítő információ). A QUD és a 3F9 kristály és dokkolt konformációi közötti RMSD 0,46 és 0,60 Å volt. Továbbá, a pterozin inhibitorok kötőhelyei összhangban voltak egy korábbi dokkolási vizsgálattal, amelyben a BACE1, AChE és BChE38 szerepelt. A vizsgálatban azonban Tetronarce californica AChE-t (PDB-kód: 1ACJ) használtak, amely a szekvenciáik eltérései miatt kissé más maradékszámokat tartalmaz, mint a humán AChE.
BACE1 dokkolás
A gátlás típusa és aktivitása alapján a (2R,3R)-pterozid C és a (2R)-pterozid B-t választották ki a vegyes típusú, illetve a nem kompetitív BACE1-gátlók dokkolási módjának bemutatására. A 3a. és b. ábra a (2R,3R)-pterozid C és a (2R)-pterozid B dokkolási modelljeit mutatja. A dokkolt vegyületek kölcsönhatásait a BACE1 aktív helyén belül a 4. ábra mutatja.

Molekuláris dokkolási modellek a vegyes típusú BACE1-inhibitor (2R,3R)-pterozid C (kék szín) és b a nem kompetitív BACE1-inhibitor (2R)-pterozid B (barna szín) számára. A dokkolt pózok a QUD röntgenkristályszerkezetére (sárga szín) vannak ráhelyezve (PDB-kód: 2WJO). A BACE1, az aktív centrum maradékai és a vegyületek szalag-, vonal- és pálcikamodellekkel vannak ábrázolva. A szaggatott vonalak színei a különböző kölcsönhatások típusait magyarázzák: hidrogénkötéses kölcsönhatások (zöld), hidrofób kölcsönhatások (rózsaszín) és π-szigma kölcsönhatások (lila). BACE1 β-site amyloid prekurzor fehérje hasító enzim 1

A QUD, b (2R,3R)-pterozid C, c (3S)-pterozid D, d (2R,3R)-pterozin C, e (2R)-pterozid B, f (2S,3R)-pterozin C és g (2R)-pterozin B ligandkölcsönhatási diagramja a BACE1 aktív helyén. A szaggatott vonalak színei a különböző kölcsönhatások típusait magyarázzák: hidrogénkötéses kölcsönhatások (zöld), hidrofób kölcsönhatások (rózsaszín) és π-szigma kölcsönhatások (lila). BACE1 β-site amyloid prekurzor fehérje hasító enzim 1
A QUD dokkolt pózában a kötési energia (B.E.) -7,59 kcal/mol volt. Amint a 4a. ábrán látható, a ligandum kinazolin gyűrűjében lévő NH2 csoport két hidrogénkötést mutatott az Asp32 és Asp228 CO csoportjaival 1,86, illetve 2,17 Å távolságban. Egy harmadik hidrogénkötés volt megfigyelhető a ligandum másik NH-csoportja és a Gly230 CO-csoportja között 2,16 Å távolságban. Leu30, Tyr71, Phe108 és Val332 közvetítették a hidrofób kölcsönhatásokat. A 4b-d ábra a (2R,3R)-pterozid C, a (3S)-pterozid D és a (2R,3R)-pterozin C (vegyes típusú BACE1-gátlók) dokkolt pózait mutatja. A Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 és Thr231 által bélelt kötőzsebben helyezkedtek el. Aktivitási szintjüket tekintve a (2R,3R)-pterozid C (IC50 = 9,74 µM), a (3S)-pterozid D (IC50 = 10,7 µM) és a (2R,3R)-pterozin C (IC50 = 26,2 µM) -7,27, -6,93 és -4,84 kcal/mol B.E.-t mutatott. A (2R,3R)-Pterozid C nagyobb hatékonyságot mutatott, mint a (2R,3R)-Pterozin C a további 2-hidroximetil-tetrahidro-pirán-3,4,5-triol csoport megléte miatt, amely három hidrogénkötést képzett (4b. ábra). A hidroximetil OH-csoportja és a tetrahidro-pirán-triolgyűrű 3-OH-csoportja két hidrogénkötést mutatott az Asp228 CO-csoportjával 2,63, illetve 2,12 Å távolságban. Ezenkívül a 3-OH csoport hidrogénkötést mutatott a Thr231-gyel 1,89 Å távolságban. A (3S)-terozid D esetében (4c. ábra) az indanongyűrűn lévő 2,2-dimetilcsoport jelenléte kissé megváltoztatta a kötési kölcsönhatásokat a (2R,3R)-terozid C-hez képest. A tetrahidro-pirán-triolgyűrű 3-OH csoportja csak egyetlen hidrogénkötést mutatott az Asp228-hoz 1,93 Å távolságban. Az indanongyűrű 3-OH csoportja azonban további hidrogénkötést mutatott az Ile126-hoz 2,34 Å távolságban. Ezek a kölcsönhatások kissé csökkentették a (3S)-terozid D aktivitását a (2R,3R)-terozid C-hez képest. A 4d. ábrán látható (2R,3R)-pterozin C dokkoló kölcsönhatásai azt mutatják, hogy az indanongyűrű 3-OH csoportja két hidrogénkötésen keresztül kötődik a Ser36-hoz és az Asn37-hez 1,83, illetve 2,07 Å távolságban. Ala39, Val69, Trp76, Ile118 és Arg128 hidrofób kölcsönhatásokban vettek részt, míg a Val69 π-szigma kölcsönhatást mutatott.
A (2R)-pterozid B, a (2S,3R)-pterozin C és a (2R)-pterozin B (nem kompetitív BACE1-gátlók) dokkolt pózai a 4e-g ábrán láthatók. Az Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 és Ile126 által bezárt üregbe dokkoltak. Aktivitási értékeiknek megfelelően a (2R)-pterozid B (IC50 = 18,0 µM), a (2S,3R)-pterozin C (IC50 = 23,1 µM) és a (2R)-pterozin B (IC50 = 29,6 µM) -6,16, -5,07 és -4,64 kcal/mol B.E.-t mutatott. Amint a 4e. ábrán látható, a (2R)-pterozid B nagyobb aktivitást mutatott, mint a (2S,3R)-pterozin C és a (2R)-pterozin B a további 2-hidroxi-metil-tetrahidro-pirán-3,4,5-triol csoport jelenlétének köszönhetően, amely négy hidrogénkötéses kölcsönhatást mutatott. A tetrahidro-pirán-triolgyűrű 4-OH csoportja és az Asn37 NH- és CO-csoportja között két hidrogénkötés volt megfigyelhető 2,48, illetve 2,19 Å távolságban. Továbbá a 3- és 5-OH csoportok további két hidrogénkötést mutattak az Ile126 és Trp76 csoportokkal 2,19, illetve 2,46 Å távolságban. A (2S,3R)-Pterozin C valamivel jobb aktivitást mutatott, mint a (2R)-Pterozin B az indanongyűrű 3. pozíciójában lévő további OH-csoport jelenlétének köszönhetően, amely 2,12 Å távolságban hidrogénkötést alakított ki a Lys107-gyel (4f. ábra). A többi kölcsönhatás hasonló volt, mint a (2R)-pterozin B esetében. Amint az a 4g. ábrán látható, a (2R)-pterozin B hidrofób kölcsönhatást mutatott a Val69, Tyr76 és Phe108 csoporttal.
AChE dokkolás
A (2R)-Pterozid B-t és a (2R,3R)-Pterozin C-t aktivitásuk és az AChE-gátlás típusa miatt a vegyes típusú, illetve a nem kompetitív AChE-gátlók dokkolási módozatainak bemutatására választottuk képviselőnek. Az 5a. és b. ábra a (2R)-pterozid B és a (2R,3R)-pterozin C dokkolási modelljeit szemlélteti. A dokkolt vegyületek AChE aktív helyén belüli kölcsönhatásait a 6. ábra mutatja.

Molekuláris dokkolási modellek a vegyes típusú AChE-inhibitor (2R)-pterozid B (kék szín) és b a nem kompetitív AChE-inhibitor (2R,3R)-pterozin C (barna szín) számára. A dokkolt pózok az E2020 röntgenkristályszerkezetére (sárga szín) vannak ráhelyezve (PDB-kód: 4EY7). Az AChE, az aktív centrum maradékai és a vegyületek szalag-, vonal- és pálcikamodellekkel vannak ábrázolva. A szaggatott vonalak színei a különböző kölcsönhatások típusait magyarázzák: hidrogénkötéses kölcsönhatások (zöld) és hidrofób kölcsönhatások (rózsaszín). AChE acetilkolinészteráz

Az a E2020, b (2R)-pterozid B, c (2R,3R)-pterozid C, d (2S,3R)-pterozin C, e (2R)-pterozin B, f (3S)-pterozid D és g (2R,3R)-pterozin C ligandkölcsönhatási diagramja az AChE aktív helyén. A szaggatott vonalak színei a különböző kölcsönhatások típusait magyarázzák: hidrogénkötéses kölcsönhatások (zöld), hidrofób kölcsönhatások (rózsaszín) és π-szigma kölcsönhatások (lila). AChE acetilkolinészteráz
Az E2020 dokkolt póza -10,28 kcal/mol B.E.-t mutatott. Ahogy a 6a. ábrán látható, az indanongyűrű CO-csoportja hidrogénkötést alakított ki a Phe295 NH-csoportjával 1,70 Å távolságban. A Trp286 és Tyr341 π-szigma kölcsönhatásokban vett részt, míg a Trp86, Trp286, Tyr337, Phe338 és Tyr341 hidrofób kölcsönhatásokat közvetített. A 6b-f ábra a (2R)-pterozid B, (2R,3R)-pterozid C, (2S,3R)-pterozin C, (2R)-pterozin B és (3S)-pterozid D (vegyes típusú AChE-gátlók) dokkolt pózait mutatja. Ezek a Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 és Tyr341 által körülvett aktív centrumban helyezkedtek el. Aktivitási értékeikkel összhangban a (2R)-pterozid B (IC50 = 2,55 µM), a (2R,3R)-pterozid C (IC50 = 3,77 µM), a (2S,3R)-pterozin C (IC50 = 12.8 µM), (2R)-pterozin B (IC50 = 16,2 µM) és (3S)-pterozid D (IC50 = 27,4 µM) -7,90, -7,49, -6,03, -5,76 és -4,91 kcal/mol B.E.-t mutatott. A (2R)-Pterozid B nagyobb hatékonyságot mutatott, mint a (2S,3R)-Pterozin C és a (2R)-Pterozin B a további 2-hidroximetil-tetrahidro-pirán-3,4,5-triol csoport jelenlétének köszönhetően, amely három hidrogénkötéses kölcsönhatást hozott létre (6b. ábra). A hidroximetil OH-csoportja és a tetrahidro-pirán-triolgyűrű 3-OH-csoportja 2,23, illetve 2,28 Å távolságban hidrogénkötést mutatott az Asn87 CO-csoportjával. Továbbá a 4-OH csoport 2,13 Å távolságban hidrogénkötést alakított ki a Trp86 CO-csoportjával. A (2R,3R)-pterozid C (6c. ábra) esetében az indanongyűrű 2. pozíciójában lévő metilcsoport nem mutatott hidrofób kölcsönhatást a Tyr341-gyel, és így viszonylag alacsonyabb aktivitást mutatott, mint a (2R)-pterozid B esetében. A Tyr341 azonban fenntartotta a hidrofób kölcsönhatást az indanongyűrű másik részével, amint az a (2R)-pterozid B esetében is látható. A (2S,3R)-pterozin C nagyobb aktivitása, mint a (2R)-pterozin B-é, annak tulajdonítható, hogy az indanongyűrű 3. pozíciójában egy további OH-csoport található, amely 1,82 Å távolságban hidrogénkötést alakított ki a Phe295-tel (6d. ábra). A többi kölcsönhatás hasonló volt a (2R)-pterozin B kölcsönhatásaihoz. Amint a 6e. ábrán látható, az indanongyűrű 6. pozíciójában lévő hidroxietilcsoport OH-csoportja hidrogénkötést alakított ki a Tyr124-gyel 2,48 Å távolságban. A Tyr124, Trp286, Tyr337, Phe338 és Tyr341 hozzájárult a hidrofób kölcsönhatásokhoz. A (2R)-pterozid B-vel és a (2R,3R)-pterozid C-vel összehasonlítva a (3S)-pterozid D az indanongyűrűn lévő 2,2-dimetilcsoport jelenléte miatt eltérő kötési kölcsönhatásokat mutatott (6f. ábra). A 2,2-dimetilcsoport jelentősen hozzájárult a (3S)-pterozid D eltérő dokkolt pózához. A tetrahidro-pirán-triol gyűrű 5-OH csoportja 1,96 Å távolságban hidrogénkötést alakított ki az Asp74-gyel. Az indanongyűrű CO-csoportja 2,97 Å távolságban hidrogénkötést mutatott a Tyr72-vel. Ezek a kölcsönhatások magyarázzák a (3S)-pterozid D alacsony aktivitását.
A (2R,3R)-pterozin C (nem kompetitív AChE-gátló) dokkolt pózát a 6g. ábra mutatja. A (2R,3R)-Pterozin C (IC50 = 23,2 µM) -5,01 kcal/mol B.E.-t mutatott. A (2R,3R)-pterozin C kötési zsebe Tyr72, Trp286, Ser293, Phe297 és Tyr341, két hidrogénkötéses kölcsönhatással. Az egyik hidrogénkötés az indanongyűrű 3-OH csoportja és a Ser293 CO-csoportja között jött létre 2,03 Å távolságban. A második hidrogénkötés az indanongyűrű 6. pozíciójában lévő hidroxietilcsoport OH-csoportja és a Tyr72 között volt megfigyelhető 1,94 Å távolságban. Az olyan rezidensek, mint a Tyr72, Trp286, Phe297 és Tyr341 hidrofób kölcsönhatásokban vettek részt.
BChE dokkolás
A BChE gátlás aktivitási szintjét és típusát figyelembe véve a (2R,3R)-pterozid C és a (2S,3R)-pterozin C-t választottuk ki a vegyes típusú, illetve nem kompetitív BChE-gátlók dokkolási módjának bemutatására. A 7a. és b. ábra a (2R,3R)-pterozid C és a (2S,3R)-pterozin C dokkolási modelljeit szemlélteti. A dokkolt vegyületek BChE aktív centrumon belüli kölcsönhatásait a 8. ábra mutatja be.
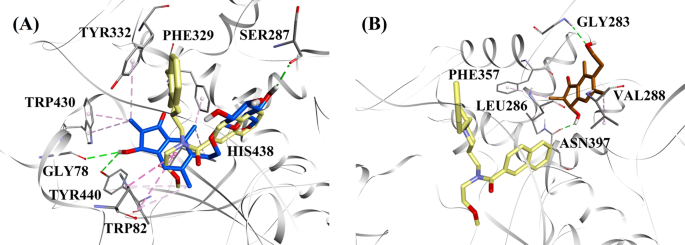
Molekuláris dokkolási modellek a vegyes típusú BChE-inhibitor (2R,3R)-pterozin C (kék szín) és b a nem kompetitív BChE-inhibitor (2S,3R)-pterozin C (barna szín) számára. A dokkolt pózok a 3F9 röntgenkristályszerkezetére (sárga szín) vannak ráhelyezve (PDB-kód: 4TPK). A BChE, az aktív centrum maradékai és a vegyületek szalag-, vonal- és pálcikamodellekkel vannak ábrázolva. A szaggatott vonalak színei a különböző kölcsönhatások típusait magyarázzák: hidrogénkötéses kölcsönhatások (zöld) és hidrofób kölcsönhatások (rózsaszín). BChE butil-kolinészteráz
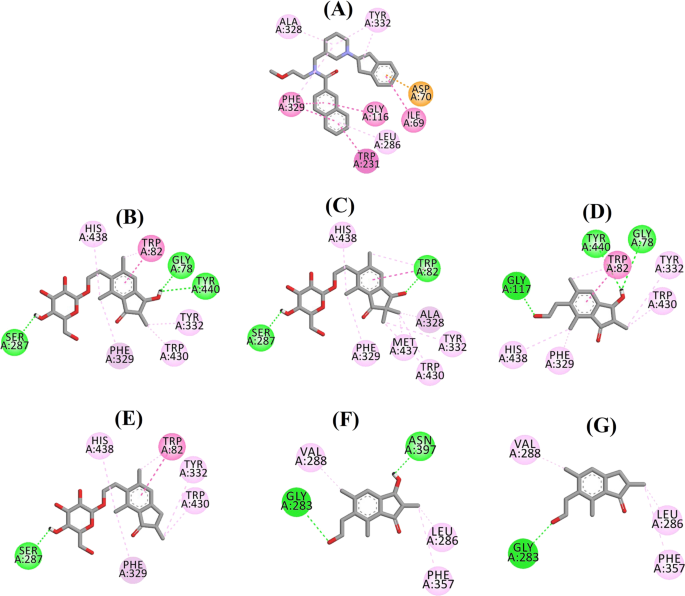
A 3F9, b (2R,3R)-pterozid C, c (3S)-pterozid D, d (2R,3R)-pterozin C, e (2R)-pterozid B, f (2S,3R)-pterozin C és g (2R)-pterozin B ligandkölcsönhatási diagramja a BChE aktív helyén. A szaggatott vonalak színei a különböző kölcsönhatások típusait magyarázzák: hidrogénkötéses kölcsönhatások (zöld), hidrofób kölcsönhatások (rózsaszín) és π-anion kölcsönhatások (arany). BChE butil-kolinészteráz
A 3F9 dokkolt póza -8,49 kcal/mol B.E.-t mutatott. Ahogy a 8a. ábrán látható, a ligandumkötésért elsősorban a hidrofób kölcsönhatások voltak felelősek. Az Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 és Tyr332 volt felelős a hidrofób kölcsönhatásokért, míg az Asp70 π-anion kölcsönhatást mutatott. A 8b-e ábra a (2R,3R)-pterozid C, a (3S)-pterozid D, a (2R,3R)-pterozin C és a (2R)-pterozid B (vegyes típusú BChE-gátlók) dokkolt pózait mutatja. Kötőzsebüket Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 és Tyr440 alkotta. Aktivitási szintjüknek megfelelően a (2R,3R)-pterozid C (IC50 = 5,29 µM), a (3S)-pterozid D (IC50 = 19,3 µM), a (2R,3R)-pterozin C (IC50 = 20,3 µM) és a (2R)-pterozid B (IC50 = 62,0 µM) -7,23, -6,59, -6,52 és -4,38 kcal/mol B.E.-t mutatott. Amint a 8b. ábrán látható, a (2R,3R)-pterozid C indanongyűrűjének 3-OH csoportja két hidrogénkötést mutatott a Gly78 és a Tyr440 csoporttal 2,87 és 2,89 Å távolságban. A (3S)-terozid D esetében az indanongyűrűn lévő 2,2-dimetilcsoport befolyásolta a 3-OH csoport kötési kölcsönhatásait (8c. ábra). A 3-OH csoport csak egy hidrogénkötést alakított ki a Trp82-vel 2,94 Å távolságban, ami a (3S)-pterozid D viszonylag alacsonyabb aktivitását eredményezte, mint a (2R,3R)-pterozid C. Ahogy a 8d. ábrán látható, a (2R,3R)-pterozin C nem tudott hidrogénkötést létrehozni a Ser287-gyel a 2-hidroximetil-tetrahidro-pirán-3,4,5-triol csoport hiánya miatt. Következésképpen alacsonyabb aktivitást mutatott, mint a (2R,3R)-pterozid C. Az indanongyűrű 3. pozíciójában lévő OH-csoport hiánya volt a felelős a (2R)-pterozid B nagyon alacsony aktivitásáért (ábra). 8e), amely a (2R,3R)-pterozid C-hez hasonlóan nem tudott hidrogénkötést képezni a Gly78 és a Tyr440-vel.
A (2S,3R)-pterozin C és a (2R)-pterozin B (nem kompetitív BChE-gátlók) dokkolt pózai a 8f, g ábrán láthatók. Ezek a dokkolt pózok a Gly283, Leu286, Val288, Phe357 és Asn397 által bezárt üregben voltak. Aktivitási szintjüket tekintve a (2S,3R)-pterozin C (IC50 = 44,3 µM) és a (2R)-pterozin B (IC50 = 48,1 µM) -5,40 és -5,06 kcal/mol B.E.-t mutatott. A (2S,3 R)-Pterozin C hatásosabb volt, mint a (2R)-Pterozin B, mivel az indanongyűrű 3. pozíciójában egy további OH-csoportot tartalmazott, amely 2,05 Å távolságban hidrogénkötést alakított ki az Asn397 CO-csoportjával (8f. ábra). Más kölcsönhatások hasonlónak bizonyultak a (2R)-pterozin B-hez. Amint a 8g. ábra mutatja, az indanongyűrű 6. pozíciójában lévő hidroxietilcsoport OH-csoportja 2,40 Å távolságban hidrogénkötést alakított ki a Gly283-mal. A Leu286, Val288 és Phe357 maradékok hidrofób kölcsönhatásokban vettek részt.
A vegyes típusú inhibitorok mind a szabad enzimhez, mind az enzim-szubsztrát komplexhez kötődnek, ami arra utal, hogy ezek a vegyületek az egyes megfelelő enzimek katalitikus helyéhez kötődhetnek. A nem kompetitív inhibitorok a szabad enzim vagy az enzim-szubsztrát komplex allosztérikus helyéhez kötődnek. Egy nemrégiben végzett vizsgálat szerint a kompetitív, vegyes típusú és nem kompetitív inhibitorok különböző helyeket foglalnak el a BACE1, az AChE és a BChE38 kötési zsebeiben. A pterozin-származékok gátlási mechanizmusának értékelésére szolgáló dokkolás során a vegyületek kötőhelyeit a gátlás típusa szerint határoztuk meg. A dokkolási eredmények azt mutatták, hogy a BACE1, AChE és BChE vegyes típusú és nem kompetitív inhibitorok kötőhelyei részben átfedik egymást az egyes megfelelő aktív helyeken, és összhangban voltak egy korábbi jelentéssel38.
BBB-permeabilitás
A PAMPA-BBB, a BBB mesterséges membránpermeabilitásának in vitro vizsgálata az egyik legmegbízhatóbb fizikai-kémiai szűrőeszköz a CNS-célzott gyógyszerek felfedezésének korai szakaszában40. A PAMPA-BBB rendszer modellezi a vegyi anyagok transzcelluláris passzív diffúzióját a BBB-n keresztül, és szigorúan passzív transzportmechanizmusokat mér egy mesterséges lipidmembránon keresztül az effektív permeabilitáson (Pe, cm/s). A BBB-permeáció előrejelzésére kialakított minta alapján a vegyületeket a következő osztályokba sorolták: (i) “CNS+” (magas BBB-permeáció előrejelzése); Pe (10-6 cm/s) > 4,00, (ii) “CNS-” (alacsony BBB-permeáció előrejelzése); Pe (10-6 cm/s) < 2,00, és (iii) “CNS+/-” (BBB-permeáció bizonytalan); Pe (10-6 cm/s) 4,00 és 2,00 között. Ennek megfelelően a (2R)-pterozin B, a (2S)-pterozin P és a (2S)-pterozin A magas BBB-permeációt mutatott 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s és 6,26 × 10-6 cm/s Pe értékekkel (4. táblázat, 4. kiegészítő információ). A (2R)-pterozin B Pe értéke 1,7-szer nagyobb volt, mint a pozitív kontrollként használt CNS-gyógyszer, a verapamilé (Pe = 34,6 × 10-6 cm/s). A (2S,3R)-Pterozin C és a (2R,3R)-Pterozin C bizonytalan BBB-permeációt mutatott 2,34-es, illetve 1,98-as Pe értékekkel. A (2R,3R)-Pterozid C, a (3S)-Pterozid D és a (2R)-Pterozid B, amelyek a vizsgált pterozin-származékok közül a legerősebb BACE1- és kolinészteráz-gátló hatást mutatták, nagyon alacsony BBB-permeabilitást mutattak. A pterozid-származékokhoz hasonlóan a 2-hidroximetil-tetrahidro-pirán-3,4,5-triol csoport megléte a BBB-permeabilitás jelentős csökkenését eredményezte. A (2R)-pterozin B-vel összehasonlítva az indanongyűrű 3. pozíciójában lévő OH-csoport további jelenléte, mint a pterozin C-ben, az indanongyűrű 2. pozíciójában lévő hidroxi-metilcsoport, mint a (2S)-pterozin A-ban, vagy az indanongyűrű 5. pozíciójában lévő hidroxi-metilcsoport, mint a (2S)-pterozin P-ben, szintén jelentősen csökkentette a BBB-permeabilitást. Figyelembe véve a kivételesen magas BBB-permeabilitást és a BACE1, AChE és BChE jelentős gátlását, a (2R)-pterozin B erős anti-AD-aktivitást mutathat.
A (2R)-pterozin B és a (2R,3R)-pterozid C hatása az Aβ peptidek neuronsejtek általi szekréciójára
A (2R)-pterozin B és a (2R,3R)-pterozid C-nek az Aβ neuronális sejtekből történő kiválasztásának csökkentésében való szerepének vizsgálatához olyan egér neuroblasztóma-sejtvonalat használtunk, amely stabilan felülreprezentálja az emberi APPswe-t. A sejtvonal az Alzheimer-kór sejtes modellje, amelyet az Aβ40 és Aβ42 túlzott szekréciója jellemez. A toxikus amiloid oligomerek az Aβ peptid két különböző hosszúságú izoformájából képződnek. Az Aβ40 a legnagyobb mennyiségben előforduló Aβ izoforma az agyban41, míg az Aβ42 jelentősen megnő az AD bizonyos formáiban42. Az Aβ40 szendvics ELISA kimutatta, hogy a (2R)-pterozin B szignifikánsan, akár 50%-kal is csökkentette a neuroblasztóma sejtekből a médiumba szekretált Aβ40 peptid mennyiségét 500 μM esetén (P < 0,01) (9a. ábra). Hasonlóképpen, az Aβ42 peptid szekréciója a neuroblastóma sejtekből szignifikánsan csökkent 500 μM (2R,3R)-pterozid C jelenlétében (P < 0,05) (9b. ábra). Összefoglalva, a (2R)-pterozin B és a (2R,3R)-pterozid C szignifikánsan csökkentette az Aβ-peptidek szekrécióját a neuroblastóma sejtekből 500 μM koncentrációban.

a A (2R)-pterozin B hatása az Aβ peptidek szekréciójára. APPswe-szekretáló neuroblasztóma sejteket 12, 60, 250 vagy 500 µM (2R)-pterozin B-vel kezeltünk 24 órán keresztül, és a kondicionált médiumokat proteáz inhibitor jelenlétében gyűjtöttük. Pozitív kontrollként β-SI-t (10 µM) használtunk. A negatív kontroll a vizsgált vegyületek hiányában tenyésztett sejteket tartalmazta. A kondicionált médiumban szekretált Aβ40 és Aβ42 mennyiségi elemzését szendvics ELISA segítségével végeztük. Három független kísérlet átlaga ± SD látható. A szekretált Aβ40 peptidek szignifikánsan csökkentek 500 µM (2R)-pterozin B jelenlétében. CON negatív kontroll, β-SI β-szekretáz inhibitor III, Aβ β-amyloid. *P < 0,05, **P < 0,01. b A (2R,3R)-pterozid C hatása az Aβ peptidek szekréciójára. Az APPswe-szekretáló neuroblastóma sejteket 12, 60, 125, 125, 250 vagy 500 µM (2R,3R)-pterozid C-vel kezeltük 24 órán keresztül, és a kondicionált médiumokat proteáz inhibitor jelenlétében gyűjtöttük. Pozitív kontrollként β-SI-t (10 µM) használtunk. A negatív kontroll a vizsgált vegyületek hiányában tenyésztett sejteket tartalmazta. A kondicionált médiumban szekretált Aβ40 és Aβ42 mennyiségi elemzését szendvics ELISA segítségével végeztük. Három független kísérlet átlaga ± SD látható. A szekretált Aβ42 peptid szignifikánsan csökkent 500 µM (2R,3R)-pterozid C jelenlétében. CON negatív kontroll, β-SI β-szekretáz inhibitor III, Aβ β-amyloid. *P < 0.05
A pterozin-származékok citotoxicitása MTT-teszt alapján43
A pterozin-származékok összességében elhanyagolható citotoxicitást mutattak különböző normál és rákos sejtvonalakkal szemben, például SH-SY5Y humán neuroblasztómával, C6 patkány gliasejtekkel, NIH3T3 egér embrió fibroblasztokkal és B16F10 egér melanomával szemben, 0 feletti LD50 értékekkel.5 mM (Kiegészítő információ 5). Különösen a (2R)-pterozin B, (3R)-pterozin D, (2S)-pterozin P, (2S)-pterozid A, (2R)-pterozid B és (2R,3R)-pterozid C nem mutatott citotoxicitást az 5 mM feletti LD50 értékkel vizsgált sejtvonalakkal szemben. Érdekes módon több pterozin relatív antiproliferatív hatást mutatott az SH-SY5Y neuronális sejtekkel szemben, szemben a mesenchymális eredetű C6 gliasejtekkel. A jelen eredmények összhangban voltak egy korábbi jelentéssel, amely szerint a pterozin-származékok nem mérgezőek az emberre26.