

A sóhíd hozzájárulása a fehérje összehajtott állapotának általános stabilitásához a mutagenezis vizsgálatokból és nukleáris mágneses rezonancia technikákból gyűjtött termodinamikai adatokkal értékelhető. Egy olyan mutált pszeudo-wild-típusú fehérje segítségével, amelyet kifejezetten a magas pH-n történő kicsapódás megakadályozására mutáltak, a sóhíd hozzájárulása a hajtogatott fehérjeállapot teljes szabad energiájához meghatározható egy pontmutáció végrehajtásával, a sóhíd megváltoztatásával és következésképpen megbontásával. Például a T4 lizozimban a 70-es maradékban lévő aszparaginsav (Asp) és a 31-es maradékban lévő hisztidin (His) között sóhidat azonosítottak (3. ábra). Aszparaginnal (Asn) végzett helyirányított mutagenezist (4. ábra) végeztünk, és három új mutánst kaptunk: Asp70Asn His31 (1. mutáns), Asp70 His31Asn (2. mutáns) és Asp70Asn His31Asn (kettős mutáns).

A mutánsok létrehozása után két módszert lehet alkalmazni a sóhídhoz kapcsolódó szabad energia kiszámítására. Az egyik módszer a vad típusú fehérje olvadási hőmérsékletének megfigyelése a három mutánséval szemben. A denaturáció a cirkuláris dichroizmus változásán keresztül követhető nyomon. Az olvadási hőmérséklet csökkenése a stabilitás csökkenését jelzi. Ezt a Becktel és Schellman által leírt módszerrel lehet számszerűsíteni, ahol a kettő közötti szabadenergia-különbséget a ΔTΔS segítségével számoljuk ki. Ezzel a számítással van néhány probléma, és csak nagyon pontos adatok esetén használható. A T4 lizozim példában a pszeudo-wild-típusú ΔS-t korábban pH 5,5-nél jelentették, így a 11 °C-os középső hőmérsékletkülönbség ennél a pH-nál megszorozva a jelentett 360 cal/(mol-K) (1,5 kJ/(mol-K)) ΔS értékkel, körülbelül -4 kcal/mol (-17 kJ/mol) szabad energiaváltozást kapunk. Ez az érték megfelel annak a szabad energiának, amellyel a sóhíd hozzájárul a fehérje stabilitásához.
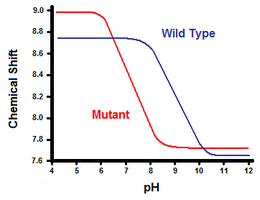
A második módszer a nukleáris mágneses rezonancia spektroszkópiát használja a sóhíd szabad energiájának kiszámításához. Titrálást végeznek, miközben rögzítik a karboxilát- vagy ammóniumcsoport melletti szén protonjainak megfelelő kémiai eltolódást. A titrálási görbe középpontja megfelel a pKa értéknek, vagyis annak a pH-értéknek, ahol a protonált:deprotonált molekulák aránya 1:1. Folytatva a T4 lizozim példáját, a 31-es hisztidin C2-protonjának eltolódását megfigyelve kapunk egy titrálási görbét (5. ábra). Az 5. ábra a titrálási görbe eltolódását mutatja a vad típus és a mutáns között, amelyben az Asp70 Asn. A kialakult sóhíd a deprotonált Asp70 és a protonált His31 között van. Ez a kölcsönhatás okozza a His31 pKa értékében megfigyelhető eltolódást. A kibontott vad típusú fehérjében, ahol a sóhíd hiányzik, a His31 pKa értéke a jelentések szerint 6,8 a mérsékelt ionerősségű H20 pufferekben. Az 5. ábra szerint a vad típus pKa értéke 9,05. Ezt a pKa-különbséget alátámasztja a His31 és az Asp70 közötti kölcsönhatás. A sóhíd fenntartása érdekében a His31 megpróbálja a protonját minél tovább megtartani. Ha a sóhíd megszakad, mint a D70N mutánsban, a pKa visszaváltozik 6,9-es értékre, ami sokkal közelebb van a His31 értékéhez a kibontott állapotban.
A pKa különbség számszerűsíthető, hogy tükrözze a sóhíd hozzájárulását a szabad energiához. A Gibbs-féle szabadenergia segítségével: ΔG = -RT ln(Keq), ahol R az egyetemes gázállandó, T a hőmérséklet kelvinben, Keq pedig az egyensúlyban lévő reakció egyensúlyi állandója. A His31 deprotonálódása savi egyensúlyi reakció, amelynek speciális Keq-je a savi disszociációs állandó, Ka: His31-H+ ⇌ His31 + H+. A pKa ezután a következőképpen függ össze a Ka-val: pKa = -log(Ka). A mutáns és a vad típus szabadenergia-különbségének kiszámítása most már elvégezhető a szabadenergia-egyenlet, a pKa definíciója, a megfigyelt pKa értékek és a természetes logaritmusok és logaritmusok közötti kapcsolat segítségével. A T4 lizozim példában ez a megközelítés egy körülbelül 3 kcal/mol-os számított hozzájárulást eredményezett a teljes szabad energiához. Hasonló megközelítést lehet alkalmazni a sóhíd másik résztvevőjével, például a T4 lizozim példában az Asp70-gyel, a His31 mutációját követő pKa eltolódásának megfigyelésével.
A megfelelő kísérlet kiválasztásakor óvatosságra int a sóhíd fehérjén belüli elhelyezkedése. A környezet nagy szerepet játszik a kölcsönhatásban. Nagy ionerősségnél a sóhíd teljesen elfedhető, mivel elektrosztatikus kölcsönhatásról van szó. A T4 lizozimban a His31-Asp70 sóhíd a fehérjén belül volt eltemetve. Az entrópia nagyobb szerepet játszik a felszíni sóhidaknál, ahol a normális esetben mozgási lehetőséggel rendelkező maradékokat az elektrosztatikus kölcsönhatás és a hidrogénkötés beszűkíti. Kimutatták, hogy ez eléggé csökkenti az entrópiát ahhoz, hogy a kölcsönhatás hozzájárulását majdnem eltörölje. A felületi sóhidak hasonlóan tanulmányozhatók, mint az eltemetett sóhidak, kettős mutációs ciklusok és NMR-titrálások alkalmazásával. Bár léteznek olyan esetek, amikor az eltemetett sóhidak hozzájárulnak a stabilitáshoz, mint bármi más, léteznek kivételek, és az eltemetett sóhidak destabilizáló hatást mutathatnak. A felszíni sóhidak bizonyos körülmények között szintén stabilizáló hatást fejthetnek ki. A stabilizáló vagy destabilizáló hatást esetről-esetre kell értékelni, és kevés általános kijelentést lehet tenni.