
Isolation and characterization of pterosin derivatives from P. aquilinum
Pterozynowe pochodne zostały wyizolowane z całych roślin P. aquilinum. W celu zbadania składników fitochemicznych z paproci słonolistnej, ekstrakt wodny był kolejno rozdzielany octanem etylu (EtOAc) i n-butanolem (n-BuOH) (Rys. 1). W wyniku wielokrotnej chromatografii kolumnowej frakcji rozpuszczalnej w EtOAc wyizolowano dziewięć pochodnych, w tym (2R)-pterozynę B (1), pterozynę Z (2), (2S)-pterozynę P (3), (3R)-pterozynę D (4), (2S)-pterozynę A (5), (2S,3R)-pterozynę C (6), (2R,3R)-pterozynę C (7), (2R)-pterozyd B (8) i pterozyd Z (9), o czystości powyżej 97% (Rys. 2). 2). W wyniku powtórnego rozdziału chromatograficznego na kolumnie frakcji rozpuszczalnej w n-BuOH wyizolowano sześć pochodnych, w tym (2S)-pterozyd A (10), (2S)-pterozyd A2 (11), (2S,3R)-pterozyd C (12), (2R,3R)-pterozyd C (13), (3S)-pterozyd D (14) i (2S)-pterozyd P (15), o czystości większej niż 97%. Struktury związków zidentyfikowano na podstawie analizy danych spektralnych, w tym MS, 1D- i 2D-NMR (Supplementary Information 1).

Struktury związków pterozyny 1-15xxx
Aktywność inhibicyjna pochodnych pterozyny wobec BACE1, AChE, i BChE
Aby ocenić potencjał anty-AD, aktywność inhibicyjną każdego związku pterozyny wobec BACE1 i cholinesteraz oceniano za pomocą odpowiednich testów inhibicyjnych in vitro (Tabela 1). Wszystkie badane pochodne pterozyny wykazywały zależną od stężenia aktywność inhibicyjną wobec BACE1 z zakresem wartości IC50 (half-maximum inhibitory concentration) wynoszącym 9,74-94,4 μM, z wyjątkiem (2S)-pterozyny A i (2S)-pterozydu P, które były nieaktywne w badanych stężeniach, w porównaniu z IC50 kwercetyny użytej jako kontrola pozytywna, która wynosiła 18,8 μM. Siła hamowania najsilniejszych inhibitorów była w kolejności: (2R,3R)-pterozyd C, (3S)-pterozyd D, (2R)-pterozyd B, (2S,3R)-pterozyd C, (2R,3R)-pterozyd C, (2S,3R)-pterozyd C oraz (2R)-pterozyd B z wartościami IC50 wynoszącymi 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 i 29,6 μM, odpowiednio. Następnie zbadaliśmy potencjał inhibicyjny pochodnych pterozyny wobec AChE. Wszystkie badane związki wykazywały znaczącą aktywność hamującą AChE, z wartościami IC50 w zakresie 2,55-110 μM, w porównaniu z IC50 wobec AChE berberyny użytej jako kontrola pozytywna, która wynosiła 0,39 μM. Związki pterozyny, które wykazywały najsilniejsze działanie hamujące AChE to (2R)-pterozyd B, (2R,3R)-pterozyd C, (2S,3R)-pterozyd C, (2S,3R)-pterozyd C i (2R)-pterozyd B z wartościami IC50 odpowiednio 2,55, 3,77, 9,17, 12,8 i 16,2 μM. Na koniec zbadaliśmy zdolności inhibicyjne pochodnych pterozyny wobec BChE. Wszystkie badane związki pterozyny wykazywały aktywność hamującą wobec BChE, z wartościami IC50, które wahały się od 5,29 do 119 μM, z wyjątkiem (3R)-pterozyny D, która była nieaktywna w badanych stężeniach, w porównaniu z IC50 berberyny wobec BChE, które wynosiło 3,32 μM. Związkami pterozynowymi, które wykazywały najsilniejsze działanie hamujące wobec BChE były (2R,3R)-pterozyd C i pterozyd Z z wartościami IC50 wynoszącymi odpowiednio 5,29 i 5,31 μM.
Zbiorczo, większość badanych pochodnych pterozyny wykazywała znaczącą aktywność hamującą wobec BACE1, AChE i BChE jednocześnie. Obecność dodatkowej grupy 2-hydroksymetylo-tetrahydro-piran-3,4,5-triolowej jak w pochodnych pterozydu znacząco zwiększała aktywność inhibicyjną wobec enzymów. Ponadto obecność dodatkowej grupy hydroksymetylowej w pozycji 2 pierścienia indanonowego (2R)-pterozyny B, jak w (2S)-pterozynie A, lub grupy metylowej, jak w (3R)-pterozynie D i pterozynie Z, zmniejszała aktywność hamującą enzymy. W szczególności obecność grupy hydroksymetylowej w pozycji 5 pierścienia indanonu, jak w (2S)-pterozynie P, zmniejszała aktywność hamującą wobec BACE1.
Parametry kinetyczne hamowania enzymów przez pochodne pterozyny
Próbując wyjaśnić sposób hamowania enzymów przez pochodne pterozyny, przeprowadziliśmy analizę kinetyczną BACE1 i cholinesteraz dla reprezentatywnych inhibitorów (Tabela 2, Informacja uzupełniająca 2). Niska wartość Ki (stała inhibicji) wskazuje na ściślejsze wiązanie enzymu i bardziej efektywny inhibitor. Ogólnie, wartości Ki związków korelowały z odpowiednimi wartościami IC50. Hamowanie BACE1 przez związki (2R,3R)-pterozyd C, (3S)-pterozyd D i (2R,3R)-pterozynę C miało charakter mieszany z wartościami Ki wynoszącymi odpowiednio 12,6, 16,5 i 27.6 µM, podczas gdy hamowanie przez (2R)-pterozyd B, (2S,3R)-pterozyd C i (2R)-pterozynę B było niekompetycyjne z wartościami Ki wynoszącymi odpowiednio 23,1, 33,8 i 38,3 µM. Hamowanie AChE przez (2R)-pterozyd B, (2R,3R)-pterozyd C, (2R)-pterozynę B, (2S,3R)-pterozynę C i (3S)-pterozyd D miało charakter mieszany z wartościami Ki wynoszącymi odpowiednio 4.89, 8,13, 12,1, 16,3 i 23,1 µM, podczas gdy (2R,3R)-pterozyna C była inhibitorem typu niekompetycyjnego z wartością Ki równą 29,6 µM. Inhibicja BChE przez (2R,3R)-pterozynę C, (2R,3R)-pterozyd C, (3S)-pterozyd D i (2R)-pterozyd B była typu mieszanego z wartościami Ki wynoszącymi odpowiednio 4,77, 9,62, 19.7, i 22,6 µM, podczas gdy (2S,3R)-pterozyd C i (2R)-pterozyd B były inhibitorami niekompetycyjnymi z wartościami Ki wynoszącymi odpowiednio 29,9 i 53,5 µM. Wyniki te sugerują, że specyficzne pochodne pterozyny mogą być skutecznymi inhibitorami BACE1, AChE i BChE.
Molekularne symulacje dokowania dla BACE1, AChE i BChE
Dostępnych jest wiele struktur krystalicznych dla BACE1 i cholinesteraz. Wybraliśmy ludzkie PDBs na podstawie struktur typu dzikiego, ligandów kokrystalicznych i rozdzielczości struktur. Do dokowania wybrano rentgenowskie struktury krystaliczne BACE1 skompleksowanej z QUD (kod PDB: 2WJO, rozdzielczość: 2,5 Å)33, AChE skompleksowanej z E2020 (kod PDB: 4EY7, rozdzielczość: 2,35 Å)34 oraz BChE skompleksowanej z 3F9 (kod PDB: 4TPK, rozdzielczość: 2,70 Å)35. Początkowo QUD, E2020 i 3F9 zostały wyodrębnione ze struktur krystalicznych i dokowane do miejsc aktywnych odpowiednio BACE1, AChE i BChE. Następnie dokowano (2R,3R)-pterozyd C, (3S)-pterozyd D, (2R,3R)-pterozyd C, (2R)-pterozyd B, (2S,3R)-pterozyd C i (2R)-pterozyd B o znanym mechanizmie inhibicji wobec BACE1, AChE i BChE w celu określenia ich SAR. Wyniki dokowania zestawiono w tabeli 3. SAR wybranych mieszanych i niekompetycyjnych inhibitorów BACE1, AChE i BChE umożliwił ocenę nowych sposobów hamowania tych enzymów przez pterozyny.
Nasz tryb dokowania E2020 był zgodny z eksperymentalnie określonym trybem wiązania zgłoszonym wcześniej z rekombinowaną ludzką AChE (rhAChE) (Informacja dodatkowa 3)34. RMSD (root-mean-square deviation) pomiędzy krystaliczną i zadokowaną konformacją E2020 wynosiło 0,54 Å, co sugeruje wiarygodność naszej konfiguracji dokowania w odtwarzaniu eksperymentalnego trybu wiązania. Ponadto, zadokowany tryb E2020 prowadził do podobnej interakcji, jak w przypadku rhAChE-E2020. W naszym badaniu cząsteczki wody zostały usunięte ze struktury krystalicznej podczas dokowania, dlatego też oddziaływania z udziałem wody nie były analizowane w niniejszej pracy. Podobnie, zadokowane tryby QUD i 3F9 były zgodne z dostępnymi danymi eksperymentalnymi dla BACE1 33 i BChE35, odpowiednio (Informacja Dodatkowa 3). RMSD pomiędzy krystalicznymi i zadokowanymi konformacjami QUD i 3F9 wynosiły odpowiednio 0,46 i 0,60 Å. Ponadto, miejsca wiązania inhibitorów pterozyny były zgodne z poprzednim badaniem dokowania, które obejmowało BACE1, AChE i BChE38. Jednakże w badaniu tym wykorzystano AChE Tetronarce californica (kod PDB: 1ACJ), która zawiera nieco inne numery reszt niż ludzka AChE z powodu różnic w ich sekwencjach.
Dokowanie BACE1
W oparciu o typ inhibicji i aktywność, (2R,3R)-pterozyd C i (2R)-pterozyd B zostały wybrane do zadokowania trybów odpowiednio mieszanego typu i niekompetycyjnych inhibitorów BACE1. Rysunek 3a, b przedstawia modele dokowania odpowiednio (2R,3R)-pterozydu C i (2R)-pterozydu B. Interakcje zadokowanych związków w miejscu aktywnym BACE1 przedstawiono na Rys. 4.

Molekularne modele dokowania dla a inhibitora BACE1 typu mieszanego (2R,3R)-pterozydu C (kolor niebieski) i b niekompetycyjnego inhibitora BACE1 (2R)-pterozydu B (kolor brązowy). Pozy zadokowane są nałożone na rentgenowską strukturę krystaliczną QUD (kolor żółty) (kod PDB: 2WJO). BACE1, reszty miejsca aktywnego i związki są przedstawione odpowiednio za pomocą modeli wstęgowych, liniowych i patykowych. Kolory linii przerywanych wyjaśniają typy różnych oddziaływań: oddziaływania wiązania wodorowego (zielony), oddziaływania hydrofobowe (różowy) i oddziaływania π-sigma (fioletowy). BACE1 β-site amyloid precursor protein cleaving enzyme 1

Schemat interakcji ligandów a QUD, b (2R,3R)-pterozyd C, c (3S)-pterozyd D, d (2R,3R)-pterozyd C, e (2R)-pterozyd B, f (2S,3R)-pterozyd C, i g (2R)-pterozyd B w miejscu aktywnym BACE1. Kolory linii przerywanych wyjaśniają typy różnych oddziaływań: oddziaływania wiązania wodorowego (zielony), oddziaływania hydrofobowe (różowy) i oddziaływania π-sigma (fioletowy). BACE1 β-site amyloid precursor protein cleaving enzyme 1
Dokowana pozycja QUD wykazywała energię wiązania (B.E.) wynoszącą -7.59 kcal/mol. Jak widać na Rys. 4a, grupa NH2 na pierścieniu chinazolinowym liganda wykazywała dwa wiązania wodorowe z grupami CO Asp32 i Asp228 w odległościach odpowiednio 1,86 i 2,17 Å. Trzecie wiązanie wodorowe zaobserwowano pomiędzy drugą grupą NH liganda a grupą CO Gly230 w odległości 2,16 Å. Leu30, Tyr71, Phe108 i Val332 pośredniczyły w oddziaływaniach hydrofobowych. Rysunek 4b-d przedstawia zadokowane pozycje odpowiednio (2R,3R)-pterozydu C, (3S)-pterozydu D i (2R,3R)-pterozyny C (inhibitory typu mieszanego BACE1). Umieszczono je w kieszeni wiążącej wyścielonej przez Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 i Thr231. Poziomy aktywności (2R,3R)-pterozydu C (IC50 = 9,74 µM), (3S)-pterozydu D (IC50 = 10,7 µM) i (2R,3R)-pterozyny C (IC50 = 26,2 µM) wynosiły odpowiednio -7,27, -6,93 i -4,84 kcal/mol. (2R,3R)-Pterozyd C wykazywał większą siłę działania niż (2R,3R)-pterozyna C ze względu na istnienie dodatkowej grupy 2-hydroksymetylo-tetrahydropiran-3,4,5-triolowej, która tworzyła trzy wiązania wodorowe (rys. 4b). Grupa OH hydroksymetylowa oraz grupa 3-OH pierścienia tetrahydropiran-triolowego wykazywały dwa wiązania wodorowe z grupą CO Asp228 w odległościach odpowiednio 2,63 i 2,12 Å. Dodatkowo grupa 3-OH wykazywała wiązanie wodorowe z Thr231 w odległości 1,89 Å. W przypadku (3S)-pterozydu D (Rys. 4c) obecność grupy 2,2-dimetylowej przy pierścieniu indanowym nieznacznie zmieniła interakcje wiążące w porównaniu z (2R,3R)-pterozydem C. Grupa 3-OH pierścienia tetrahydropiran-triolu wykazywała tylko pojedyncze wiązanie wodorowe z Asp228 w odległości 1,93 Å. Natomiast grupa 3-OH pierścienia indanonowego wykazywała dodatkowe wiązanie wodorowe z Ile126 w odległości 2,34 Å. Oddziaływania te nieznacznie obniżały aktywność (3S)-pterozydu D w porównaniu z (2R,3R)-pterozydem C. Interakcje dokowania (2R,3R)-pterozydu C przedstawione na Rys. 4d pokazują, że grupa 3-OH pierścienia indanonowego wiąże się z Ser36 i Asn37 poprzez dwa wiązania wodorowe w odległościach odpowiednio 1,83 i 2,07 Å. Ala39, Val69, Trp76, Ile118 i Arg128 były zaangażowane w oddziaływania hydrofobowe, podczas gdy Val69 wykazywał oddziaływanie π-sigma.
Pozycje dokowane (2R)-pterozydu B, (2S,3R)-pterozyny C i (2R)-pterozyny B (niekompetycyjnych inhibitorów BACE1) pokazano odpowiednio na Rys. 4e-g. Zostały one zadokowane do wnęki zamkniętej przez Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 i Ile126. Zgodnie z ich wartościami aktywności, (2R)-pterozyd B (IC50 = 18,0 µM), (2S,3R)-pterozyd C (IC50 = 23,1 µM) i (2R)-pterozyd B (IC50 = 29,6 µM) wykazywały B.E. wynoszące odpowiednio -6,16, -5,07 i -4,64 kcal/mol. Jak pokazano na Rys. 4e, (2R)-pterozyd B wykazywał wyższą aktywność niż (2S,3R)-pterozyna C i (2R)-pterozyna B ze względu na obecność dodatkowej grupy 2-hydroksymetylo-tetrahydro-piran-3,4,5-triolowej, która wykazywała cztery oddziaływania w postaci wiązań wodorowych. Dwa wiązania wodorowe zaobserwowano pomiędzy grupą 4-OH pierścienia tetrahydropiran-triolowego a grupami NH i CO Asn37 w odległościach odpowiednio 2,48 i 2,19 Å. Ponadto, grupy 3- i 5-OH wykazywały dwa dodatkowe wiązania wodorowe z Ile126 i Trp76 w odległościach odpowiednio 2,19 i 2,46 Å. (2S,3R)-Pterozyna C wykazywała nieco lepszą aktywność niż (2R)-pterozyna B ze względu na obecność dodatkowej grupy OH w pozycji 3 pierścienia indanonowego, która tworzyła wiązanie wodorowe z Lys107 w odległości 2,12 Å (Rys. 4f). Pozostałe oddziaływania były podobne jak w przypadku (2R)-pterozyny B. Jak widać na Rys. 4g, (2R)-pterozyna B wykazywała oddziaływania hydrofobowe z Val69, Tyr76 i Phe108.
Dokowanie AChE
(2R)-Pterozyd B i (2R,3R)-pterozyna C zostały wybrane jako przedstawiciele do zadokowania trybów odpowiednio mieszanych i niekompetycyjnych inhibitorów AChE, ze względu na ich aktywność i typ inhibicji AChE. Rysunek 5a, b ilustruje modele dokowania (2R)-pterozydu B i (2R,3R)-pterozyny C, odpowiednio. Interakcje zadokowanych związków w miejscu aktywnym AChE przedstawiono na Rys. 6.

Molekularne modele dokowania dla a inhibitora AChE typu mieszanego (2R)-pterozydu B (kolor niebieski) i b niekompetycyjnego inhibitora AChE (2R,3R)-pterozydu C (kolor brązowy). Pozy zadokowane są nałożone na rentgenowską strukturę krystaliczną E2020 (kolor żółty) (kod PDB: 4EY7). AChE, reszty miejsca aktywnego i związki są przedstawione odpowiednio za pomocą modeli wstęgowych, liniowych i patykowych. Kolory linii przerywanych wyjaśniają typy różnych oddziaływań: oddziaływania wiązania wodorowego (zielony) i oddziaływania hydrofobowe (różowy). AChE acetylocholinoesteraza

Schemat interakcji ligandów a E2020, b (2R)-pterozydu B, c (2R,3R)-pterozydu C, d (2S,3R)-pterozynu C, e (2R)-pterozynu B, f (3S)-pterozydu D i g (2R,3R)-pterozynu C w miejscu aktywnym AChE. Kolory linii przerywanych wyjaśniają rodzaje poszczególnych oddziaływań: oddziaływania typu wiązanie wodorowe (zielony), oddziaływania hydrofobowe (różowy) i oddziaływania typu π-sigma (fioletowy). AChE acetylocholinoesteraza
Dokowana pozycja E2020 wykazała B.E. równy -10.28 kcal/mol. Jak pokazano na Rys. 6a, grupa CO pierścienia indanonowego tworzyła wiązanie wodorowe z grupą NH w Phe295 w odległości 1,70 Å. Trp286 i Tyr341 uczestniczyły w oddziaływaniach π-sigma, podczas gdy Trp86, Trp286, Tyr337, Phe338 i Tyr341 pośredniczyły w oddziaływaniach hydrofobowych. Na rysunkach 6b-f przedstawiono dokowane pozy odpowiednio (2R)-pterozydu B, (2R,3R)-pterozydu C, (2S,3R)-pterozynu C, (2R)-pterozynu B i (3S)-pterozydu D (inhibitory AChE typu mieszanego). Zostały one umieszczone w miejscu aktywnym w otoczeniu Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 i Tyr341. Zgodnie z ich wartościami aktywności, (2R)-pterozyd B (IC50 = 2,55 µM), (2R,3R)-pterozyd C (IC50 = 3,77 µM), (2S,3R)-pterozyd C (IC50 = 12.8 µM), (2R)-pterozyna B (IC50 = 16,2 µM) i (3S)-pterozyd D (IC50 = 27,4 µM) wykazywały B.E. odpowiednio -7,90, -7,49, -6,03, -5,76 i -4,91 kcal/mol. (2R)-Pterozyd B wykazywał wyższą siłę działania niż (2S,3R)-pterozyna C i (2R)-pterozyna B ze względu na obecność dodatkowej grupy 2-hydroksymetylo-tetrahydro-piran-3,4,5-triolowej, która tworzyła trzy wiązania wodorowe (ryc. 6b). Grupa OH hydroksymetylowa oraz grupa 3-OH pierścienia tetrahydropiran-triolu wykazywały wiązania wodorowe z grupą CO Asn87 w odległościach odpowiednio 2,23 i 2,28 Å. Ponadto, grupa 4-OH tworzyła wiązanie wodorowe z grupą CO Trp86 w odległości 2,13 Å. W przypadku (2R,3R)-pterozydu C (Rys. 6c) grupa metylowa w pozycji 2 pierścienia indanonowego nie wykazywała oddziaływania hydrofobowego z Tyr341 i w związku z tym wykazywała stosunkowo mniejszą aktywność niż (2R)-pterozyd B. Jednak Tyr341 utrzymywał oddziaływanie hydrofobowe z Tyr341. Jednakże Tyr341 utrzymywał oddziaływanie hydrofobowe z drugą częścią pierścienia indanonowego, jak wykazano w przypadku (2R)-pterozydu B. Wyższą aktywność (2S,3R)-pterozyny C niż (2R)-pterozydu B przypisano istnieniu dodatkowej grupy OH w pozycji-3 pierścienia indanonowego, która tworzyła wiązanie wodorowe z Phe295 w odległości 1,82 Å (Rys. 6d). Pozostałe oddziaływania były porównywalne z oddziaływaniami (2R)-pterozyny B. Jak pokazano na Rys. 6e, grupa OH grupy hydroksyetylowej w pozycji 6 pierścienia indanonowego tworzyła wiązanie wodorowe z Tyr124 w odległości 2,48 Å. Tyr124, Trp286, Tyr337, Phe338 i Tyr341 przyczyniły się do powstania oddziaływań hydrofobowych. W porównaniu z (2R)-pterozydem B i (2R,3R)-pterozydem C, (3S)-pterozyd D wykazywał odmienne oddziaływania wiążące ze względu na obecność grupy 2,2-dimetylowej przy pierścieniu indanonowym (Rys. 6f). Grupa 2,2-dimetylowa w znacznym stopniu przyczyniła się do odmiennej pozycji dokowanej (3S)-pterozydu D. Grupa 5-OH pierścienia tetrahydropiran-triolu tworzyła wiązanie wodorowe z Asp74 w odległości 1,96 Å. Grupa CO pierścienia indanonowego tworzyła wiązanie wodorowe z Tyr72 w odległości 2,97 Å. Te oddziaływania odpowiadają za niską aktywność (3S)-pterozydu D.
Pozę zadokowaną (2R,3R)-pterozyny C (niekompetycyjnego inhibitora AChE) przedstawiono na Rys. 6g. (2R,3R)-Pterozyna C (IC50 = 23,2 µM) wykazała B.E. równą -5,01 kcal/mol. Kieszeń wiążąca (2R,3R)-pterozyny C obejmowała Tyr72, Trp286, Ser293, Phe297 i Tyr341, z dwoma oddziaływaniami w postaci wiązań wodorowych. Jedno z wiązań wodorowych powstało pomiędzy grupą 3-OH pierścienia indanonowego a grupą CO Ser293 w odległości 2,03 Å. Drugie wiązanie wodorowe zaobserwowano pomiędzy grupą OH grupy hydroksyetylowej obecnej w pozycji 6 pierścienia indanonowego a Tyr72 w odległości 1,94 Å. Rezydy takie jak Tyr72, Trp286, Phe297 i Tyr341 uczestniczyły w oddziaływaniach hydrofobowych.
Dokowanie BChE
Rozważając poziomy aktywności i typ inhibicji BChE, (2R,3R)-pterozyd C i (2S,3R)-pterozyd C zostały wybrane do zademonstrowania zadokowanych trybów odpowiednio mieszanych i niekompetycyjnych inhibitorów BChE. Rysunek 7a, b ilustruje modele dokowania (2R,3R)-pterozydu C i (2S,3R)-pterozyny C, odpowiednio. Interakcje zadokowanych związków wewnątrz miejsca aktywnego BChE przedstawiono na Rys. 8.
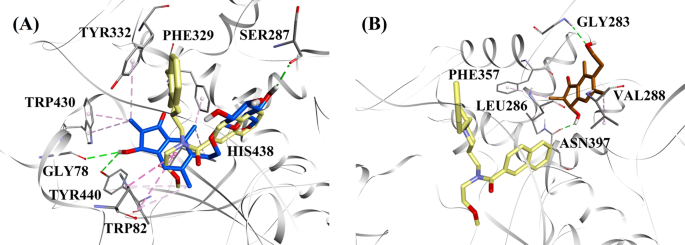
Molekularne modele dokowania dla a inhibitora BChE typu mieszanego (2R,3R)-pterozydu C (kolor niebieski) i b niekompetycyjnego inhibitora BChE (2S,3R)-pterozydu C (kolor brązowy). Pozy zadokowane są nałożone na rentgenowską strukturę krystaliczną 3F9 (kolor żółty) (kod PDB: 4TPK). BChE, reszty miejsca aktywnego i związki są przedstawione odpowiednio za pomocą modeli wstęgowych, liniowych i patykowych. Kolory linii przerywanych oznaczają rodzaje oddziaływań: wiązania wodorowe (zielony) i oddziaływania hydrofobowe (różowy). BChE butyrylocholinoesteraza
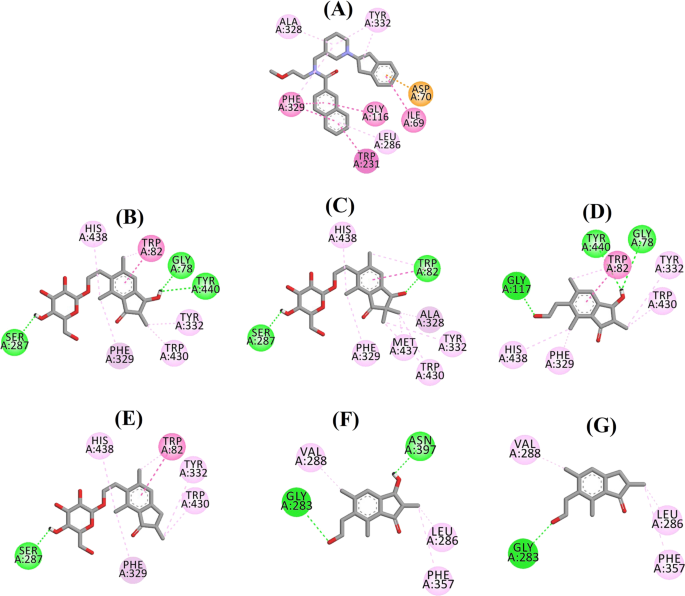
Schemat interakcji ligandów a 3F9, b (2R,3R)-pterozydu C, c (3S)-pterozydu D, d (2R,3R)-pterozynu C, e (2R)-pterozydu B, f (2S,3R)-pterozynu C i g (2R)-pterozynu B w miejscu aktywnym BChE. Kolory linii przerywanych wyjaśniają rodzaje poszczególnych oddziaływań: oddziaływania wiązania wodorowego (kolor zielony), oddziaływania hydrofobowe (kolor różowy) oraz oddziaływania π-anionowe (kolor złoty). BChE butyrylocholinoesteraza
Dokowana pozycja 3F9 wykazała B.E. równy -8.49 kcal/mol. Jak widać na Rys. 8a, za wiązanie ligandu odpowiedzialne były głównie oddziaływania hydrofobowe. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 i Tyr332 odpowiadały za oddziaływania hydrofobowe, podczas gdy Asp70 wykazywał oddziaływanie π-anionowe. Rysunki 8b-e ilustrują dokowane pozy odpowiednio (2R,3R)-pterozydu C, (3S)-pterozydu D, (2R,3R)-pterozyny C i (2R)-pterozydu B (inhibitory BChE typu mieszanego). Ich kieszeń wiążąca składała się z Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 i Tyr440. Zgodnie z ich poziomami aktywności, (2R,3R)-pterozyd C (IC50 = 5,29 µM), (3S)-pterozyd D (IC50 = 19,3 µM), (2R,3R)-pterozyd C (IC50 = 20,3 µM) i (2R)-pterozyd B (IC50 = 62,0 µM) wykazywały B.E. odpowiednio -7,23, -6,59, -6,52 i -4,38 kcal/mol. Jak pokazano na Rys. 8b, grupa 3-OH pierścienia indanonowego (2R,3R)-pterozydu C wykazywała dwa wiązania wodorowe z Gly78 i Tyr440 w odległościach odpowiednio 2,87 i 2,89 Å. W przypadku (3S)-pterozydu D grupa 2,2-dimetylowa przy pierścieniu indanonowym wpływała na oddziaływania wiążące grupy 3-OH (Rys. 8c). Grupa 3-OH utworzyła tylko jedno wiązanie wodorowe z Trp82 w odległości 2,94 Å, co spowodowało stosunkowo mniejszą aktywność (3S)-pterozydu D niż (2R,3R)-pterozydu C. Jak pokazano na Rys. 8d, (2R,3R)-pterozyd C nie wytworzył wiązania wodorowego z Ser287 z powodu braku grupy 2-hydroksymetylo-tetrahydro-piran-3,4,5-triolowej. W związku z tym jego aktywność była niższa niż (2R,3R)-pterozydu C. Brak grupy OH w pozycji 3 pierścienia indanonowego okazał się odpowiedzialny za bardzo niską aktywność (2R)-pterozydu B (ryc. 8e). 8e), który nie tworzył wiązań wodorowych z Gly78 i Tyr440 podobnie jak (2R,3R)-pterozyd C.
Pozycje dokowane (2S,3R)-pterozydu C i (2R)-pterozydu B (niekompetycyjnych inhibitorów BChE) przedstawiono odpowiednio na Rys. 8f, g. Te zadokowane pozy były zawarte we wnęce zamkniętej przez Gly283, Leu286, Val288, Phe357 i Asn397. Aktywność (2S,3R)-pterozyny C (IC50 = 44,3 µM) i (2R)-pterozyny B (IC50 = 48,1 µM) wynosiła B.E. odpowiednio -5,40 i -5,06 kcal/mol. (2S,3 R)-Pterozyna C była silniejsza od (2R)-pterozyny B ze względu na obecność dodatkowej grupy OH w pozycji 3 pierścienia indanonowego, która tworzyła wiązanie wodorowe z grupą CO w Asn397 w odległości 2,05 Å (Rys. 8f). Inne oddziaływania okazały się podobne jak w przypadku (2R)-pterozyny B. Jak widać na Rys. 8g, grupa OH w grupie hydroksyetylowej w pozycji 6 pierścienia indanonowego tworzyła wiązanie wodorowe z Gly283 w odległości 2,40 Å. Reszty Leu286, Val288 i Phe357 uczestniczyły w oddziaływaniach hydrofobowych.
Inhibitory typu mieszanego wiążą się zarówno z wolnym enzymem, jak i kompleksem enzym-substrat, co wskazuje, że związki te mogą wiązać się z miejscem katalitycznym każdego z odpowiadających im enzymów. Inhibitory niekompetycyjne wiążą się z miejscem allosterycznym wolnego enzymu lub kompleksu enzym-substrat. Ostatnie badania sugerują, że inhibitory kompetycyjne, mieszane i niekompetycyjne zajmują różne miejsca w kieszeniach wiążących BACE1, AChE i BChE38. Podczas dokowania w celu oceny mechanizmu hamowania pochodnych pterozyny, miejsca wiązania związków zostały określone zgodnie z ich typem inhibicji. Wyniki dokowania wskazały, że miejsca wiązania inhibitorów typu mieszanego i niekompetycyjnych dla BACE1, AChE i BChE częściowo nakładają się na siebie w każdym odpowiadającym im miejscu aktywnym i były zgodne z wcześniejszym doniesieniem38.
Przepuszczalność BBB
PAMPA-BBB, badanie przepuszczalności sztucznej błony BBB in vitro, jest jednym z najbardziej wiarygodnych fizykochemicznych narzędzi przesiewowych we wczesnym etapie odkrywania leków skierowanych do OUN40. System PAMPA-BB modeluje przezkomórkową bierną dyfuzję substancji chemicznych przez BBB i mierzy mechanizmy ściśle biernego transportu przez sztuczną błonę lipidową na podstawie efektywnej przepuszczalności (Pe, cm/s). Na podstawie wzorca ustalonego dla przewidywania przenikania przez BBB, związki sklasyfikowano jako (i) „CNS+” (przewidywane wysokie przenikanie przez BBB); Pe (10-6 cm/s) > 4,00, (ii) „CNS-” (przewidywane niskie przenikanie przez BBB); Pe (10-6 cm/s) < 2,00 oraz (iii) „CNS+/-” (niepewne przenikanie przez BBB); Pe (10-6 cm/s) od 4,00 do 2,00. Odpowiednio, (2R)-pterozyna B, (2S)-pterozyna P i (2S)-pterozyna A wykazywały wysokie przenikanie przez BBB z wartościami Pe wynoszącymi odpowiednio 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s i 6,26 × 10-6 cm/s (Tabela 4, Informacja dodatkowa 4). Wartość Pe (2R)-pterozyny B była 1,7-krotnie wyższa niż leku działającego na OUN – werapamilu (Pe = 34,6 × 10-6 cm/s), który został użyty jako kontrola pozytywna. (2S,3R)-Pterozyna C i (2R,3R)-pterozyna C wykazywały niepewne przenikanie przez BBB z wartościami Pe wynoszącymi odpowiednio 2,34 i 1,98. (2R,3R)-pterozyd C, (3S)-pterozyd D i (2R)-pterozyd B, które wykazywały najsilniejsze działanie hamujące BACE1- i cholinoesterazę spośród badanych pochodnych pterozyny, wykazywały bardzo niską przenikalność do BBB. Obecność grupy 2-hydroksymetylo-tetrahydropiran-3,4,5-triolowej, tak jak w pochodnych pterozydu, spowodowała znaczne zmniejszenie przepuszczalności BBB. W porównaniu z (2R)-pterozyną B, dodatkowa obecność grupy OH w pozycji 3 pierścienia indanonowego, jak w pterozynie C, grupy hydroksymetylowej w pozycji 2 pierścienia indanonowego, jak w (2S)-pterozynie A lub grupy hydroksymetylowej w pozycji 5 pierścienia indanonowego, jak w (2S)-pterozynie P, również znacząco zmniejszała przepuszczalność BBB. Biorąc pod uwagę wyjątkowo wysoką przepuszczalność BBB i znaczące hamowanie BACE1, AChE i BChE, (2R)-pterozyna B może potencjalnie wykazywać silną aktywność anty-AD.
Wpływ (2R)-pterozyny B i (2R,3R)-pterozydu C na wydzielanie peptydów Aβ przez komórki neuronalne
Badanie funkcji (2R)-pterozyny B i (2R,3R)-pterozydu C w zmniejszaniu wydalania Aβ z komórek neuronalnych, użyliśmy linii komórkowej mysiego neuroblastoma, która trwale nadekspresjonowała ludzkie APPswe. Linia komórkowa jest komórkowym modelem AD charakteryzującym się nadmiernym wydzielaniem Aβ40 i Aβ42. Toksyczne oligomery amyloidu powstają z dwóch izoform peptydu Aβ o różnej długości. Aβ40 jest najobficiej występującą izoformą Aβ w mózgu41, podczas gdy Aβ42 znacznie wzrasta w niektórych postaciach choroby Alzheimera42. Sandwich ELISA dla Aβ40 wykazała, że (2R)-pterozyna B znacząco zmniejszyła ilość peptydu Aβ40 wydzielanego z komórek neuroblastoma do mediów do 50% przy 500 μM (P < 0,01) (ryc. 9a). Podobnie, wydzielanie peptydu Aβ42 przez komórki neuroblastoma istotnie zmniejszyło się w obecności 500 μM (2R,3R)-pterozydu C (P < 0,05) (ryc. 9b). Podsumowując, (2R)-pterozyna B i (2R,3R)-pterozyd C znacząco zmniejszały wydzielanie peptydów Aβ z komórek neuroblastoma w stężeniu 500 μM.

a Wpływ (2R)-pterozyny B na wydzielanie peptydów Aβ. Komórki neuroblastoma wydzielające APPswe poddawano działaniu 12, 60, 250 lub 500 µM (2R)-pterozyny B przez 24 h, a media kondycjonowane zbierano w obecności inhibitora proteazy. Jako kontrolę pozytywną stosowano β-SI (10 µM). Kontrolę negatywną stanowiły komórki hodowane w nieobecności badanych związków. Analizę ilościową wydzielanych Aβ40 i Aβ42 w mediach kondycjonowanych przeprowadzono metodą ELISA typu sandwich. Przedstawiono średnie ± SD z trzech niezależnych eksperymentów. Wydzielane peptydy Aβ40 ulegały znacznemu zmniejszeniu w obecności 500 µM (2R)-pterozyny B. CON kontrola negatywna, β-SI β-sekretaza inhibitor III, Aβ β-amyloid. *P < 0,05, **P < 0,01. b Wpływ (2R,3R)-pterozydu C na wydzielanie peptydów Aβ. Komórki neuroblastoma wydzielające APPswe poddawano działaniu 12, 60, 125, 250 lub 500 µM (2R,3R)-pterozydu C przez 24 h, a media kondycjonowane zbierano w obecności inhibitora proteazy. Jako kontrolę pozytywną stosowano β-SI (10 µM). Kontrolę ujemną stanowiły komórki hodowane w nieobecności badanych związków. Analizę ilościową wydzielanych Aβ40 i Aβ42 w podłożu kondycjonowanym przeprowadzono metodą ELISA typu sandwich. Przedstawiono średnie ± SD z trzech niezależnych eksperymentów. Wydzielany peptyd Aβ42 znacząco zmniejszał się w obecności 500 µM (2R,3R)-pterozydu C. CON kontrola negatywna, β-SI β-sekretaza inhibitor III, Aβ β-amyloid. *P < 0.05
Cytotoksyczność pochodnych pterozyny na podstawie testu MTT43
Ogółem, pochodne pterozyny wykazywały nieistotną cytotoksyczność wobec różnych normalnych i nowotworowych linii komórkowych, takich jak ludzki neuroblastoma SH-SY5Y, szczurze komórki glejowe C6, fibroblasty NIH3T3 z zarodka myszy i czerniak B16F10 myszy z wartościami LD50 powyżej 0.5 mM (Informacja dodatkowa 5). W szczególności (2R)-pterozyna B, (3R)-pterozyna D, (2S)-pterozyna P, (2S)-pterozyd A, (2R)-pterozyd B i (2R,3R)-pterozyd C nie wykazywały cytotoksyczności wobec badanych linii komórkowych z wartościami LD50 powyżej 5 mM. Co intrygujące, kilka pterozyn wykazało względne działanie antyproliferacyjne wobec komórek neuronalnych SH-SY5Y w porównaniu z komórkami glejowymi C6 pochodzenia mezenchymalnego. Obecne wyniki były zgodne z poprzednim raportem, który sugerował, że pochodne pterozyny są nietoksyczne dla ludzi26.
.