

Wkład mostka solnego w ogólną stabilność do stanu złożonego białka może być oceniony poprzez dane termodynamiczne zebrane z badań mutagenezy i technik magnetycznego rezonansu jądrowego. Wykorzystując zmutowane białko typu pseudo-wild specjalnie zmutowane w celu zapobiegania wytrącaniu przy wysokim pH, udział mostka solnego w ogólnej energii swobodnej stanu sfałdowanego białka może być określony przez wykonanie mutacji punktowej, zmieniającej i, w konsekwencji, przerywającej mostek solny. Na przykład, w lizozymie T4 zidentyfikowano istnienie mostka solnego pomiędzy kwasem asparaginowym (Asp) w reszcie 70 i histydyną (His) w reszcie 31 (Rysunek 3). Przeprowadzono mutagenezę ukierunkowaną miejscowo z asparaginą (Asn) (Rysunek 4) uzyskując trzy nowe mutanty: Asp70Asn His31 (Mutant 1), Asp70 His31Asn (Mutant 2) oraz Asp70Asn His31Asn (Mutant podwójny).

Po ustaleniu mutantów można zastosować dwie metody obliczania energii swobodnej związanej z mostkiem solnym. Jedna metoda obejmuje obserwację temperatury topnienia białka typu dzikiego w porównaniu z temperaturą topnienia trzech mutantów. Denaturacja może być monitorowana poprzez zmianę w dichroizmie kołowym. Obniżenie temperatury topnienia wskazuje na obniżenie stabilności. Jest to określane ilościowo za pomocą metody opisanej przez Becktela i Schellmana, gdzie różnica energii swobodnej między nimi jest obliczana jako ΔTΔS. Z obliczeniami tymi wiążą się pewne problemy i mogą być one stosowane tylko przy bardzo dokładnych danych. W przykładzie lizozymu T4, ΔS typu pseudowilgotnego została wcześniej podana przy pH 5,5, więc różnica temperatury w punkcie środkowym wynosząca 11°C przy tym pH pomnożona przez podaną ΔS wynoszącą 360 kal/(mol-K) (1,5 kJ/(mol-K)) daje zmianę energii swobodnej wynoszącą około -4 kcal/mol (-17 kJ/mol). Wartość ta odpowiada ilości energii swobodnej wnoszonej do stabilności białka przez mostek solny.
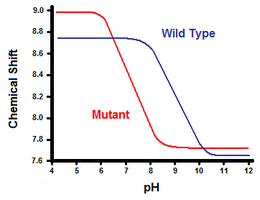
Druga metoda wykorzystuje spektroskopię magnetycznego rezonansu jądrowego do obliczenia energii swobodnej mostka solnego. Wykonuje się miareczkowanie, rejestrując jednocześnie przesunięcie chemiczne odpowiadające protonom węgla przylegającego do grupy karboksylanowej lub amonowej. Punkt środkowy krzywej miareczkowania odpowiada pKa, czyli pH, w którym stosunek cząsteczek protonowanych do zdeprotonowanych wynosi 1:1. Kontynuując przykład lizozymu T4, krzywą miareczkowania uzyskuje się poprzez obserwację przesunięcia protonu C2 histydyny 31 (Rysunek 5). Rysunek 5 przedstawia przesunięcie krzywej miareczkowania pomiędzy typem dzikim a mutantem, w którym Asp70 jest Asn. Mostek solny tworzy się pomiędzy zdeprotonowanym Asp70 i protonowaną His31. Ta interakcja powoduje przesunięcie widoczne w pKa His31. W rozwiniętym białku typu dzikiego, gdzie mostek solny jest nieobecny, His31 ma pKa równe 6,8 w buforach H20 o umiarkowanej sile jonowej. Rysunek 5 pokazuje pKa typu dzikiego równe 9,05. Ta różnica w pKa jest poparta interakcją His31 z Asp70. Aby utrzymać mostek solny, His31 będzie starał się utrzymać swój proton tak długo, jak to możliwe. Kiedy mostek solny jest zaburzony, jak w mutancie D70N, pKa przesuwa się z powrotem do wartości 6,9, znacznie bliżej wartości His31 w stanie rozwiniętym.
Różnica w pKa może być określona ilościowo, aby odzwierciedlić wkład mostka solnego do energii swobodnej. Używając wolnej energii Gibbsa:ΔG = -RT ln(Keq), gdzie R jest uniwersalną stałą gazową, T jest temperaturą w kelwinach, a Keq jest stałą równowagi reakcji w stanie równowagi. Deprotonacja His31 jest reakcją równowagi kwasów ze specjalną stałą Keq znaną jako stała dysocjacji kwasów, Ka: His31-H+ ⇌ His31 + H+. Wartość pKa jest więc związana z Ka następującym wzorem: pKa = -log(Ka). Obliczenie różnicy energii swobodnej mutanta i dzikiego typu może być teraz wykonane z wykorzystaniem równania energii swobodnej, definicji pKa, obserwowanych wartości pKa oraz zależności pomiędzy logarytmami naturalnymi i logarytmami. W przykładzie lizozymu T4, takie podejście dało wyliczony wkład około 3 kcal/mol do całkowitej energii swobodnej. Podobne podejście można przyjąć w przypadku innego uczestnika mostka solnego, takiego jak Asp70 w przykładzie lizozymu T4, monitorując jego przesunięcie w pKa po mutacji His31.
Uwaga przy wyborze odpowiedniego eksperymentu dotyczy lokalizacji mostka solnego w obrębie białka. Środowisko odgrywa dużą rolę w interakcji. Przy wysokich siłach jonowych, mostek solny może być całkowicie zamaskowany, ponieważ oddziaływanie elektrostatyczne jest zaangażowane. Mostek solny His31-Asp70 w lizozymie T4 był zakopany wewnątrz białka. Entropia odgrywa większą rolę w powierzchniowych mostkach solnych, gdzie reszty, które normalnie mają możliwość ruchu, są ograniczone przez oddziaływania elektrostatyczne i wiązania wodorowe. Wykazano, że zmniejsza to entropię na tyle, że wkład oddziaływań jest niemalże wymazywany. Powierzchniowe mostki solne mogą być badane podobnie jak pogrzebane mostki solne, z wykorzystaniem cykli podwójnych mutantów i miareczkowania NMR. Chociaż istnieją przypadki, w których zakopane mostki solne przyczyniają się do stabilności, jak wszystko inne, istnieją wyjątki i zakopane mostki solne mogą wykazywać działanie destabilizujące. Również powierzchniowe mostki solne, w pewnych warunkach, mogą wykazywać efekt stabilizujący. Efekt stabilizujący lub destabilizujący musi być oceniany dla każdego przypadku osobno i niewiele można powiedzieć na ten temat.
.