Inhibarea enzimei 1 de clivare a proteinei precursoare a amiloidului β-site și a colinesterazelor de către pterosine printr-o relație specifică structură-activitate cu o puternică permeabilitate BBB
Isolarea și caracterizarea derivaților de pterosină din P. aquilinum
Derivații de pterosină au fost izolați din plantele întregi de P. aquilinum. Pentru investigarea constituenților fitochimici din feriga de fagure, un extract de apă a fost partiționat succesiv cu acetat de etil (EtOAc) și n-butanol (n-BuOH) (Fig. 1). Cromatografia pe coloană repetată a fracțiunii solubile în EtOAc a dus la izolarea a nouă derivați, inclusiv (2R)-pterosina B (1), pterosina Z (2), (2S)-pterosina P (3), (3R)-pterosin D (4), (2S)-pterosin A (5), (2S,3R)-pterosin C (6), (2R,3R)-pterosin C (7), (2R)-pteroside B (8) și pteroside Z (9), cu purități mai mari de 97 % (Fig. 2). Separarea cromatografică repetată pe coloană a fracțiunii solubile în n-BuOH a dus la izolarea a șase derivați, inclusiv (2S)-pteroside A (10), (2S)-pteroside A2 (11), (2S,3R)-pteroside C (12), (2R,3R)-pteroside C (13), (3S)-pteroside D (14) și (2S)-pteroside P (15), cu purități mai mari de 97%. Structurile compușilor au fost identificate prin analiza datelor spectrale, inclusiv MS, 1D- și 2D-NMR (Informații suplimentare 1).
Fig. 2
Structurile compușilor de pterosină 1-15xxx
Activitatea inhibitorie a derivaților de pterosină împotriva BACE1, AChE, și BChE
Pentru a evalua potențialul anti-AD, activitatea inhibitorie a fiecărui compus de pterosin împotriva BACE1 și a colinesterazelor a fost evaluată prin testele de inhibiție in vitro respective (tabelul 1). Toți derivații de pterosină testați au prezentat activități inhibitoare dependente de concentrație împotriva BACE1, cu un interval de valori IC50 (concentrația inhibitorie semimaximă) de 9,74-94,4 μM, cu excepția (2S)-pterosinei A și a (2S)-pterosidei P, care au fost inactive la concentrațiile testate, în comparație cu IC50 a quercetinei utilizate ca martor pozitiv, care a fost de 18,8 μM. Puterea inhibitorie a celor mai puternici inhibitori a fost în ordinea (2R,3R)-pterosidei C, (3S)-pterosidei D, (2R)-pterosidei B, (2S,3R)-pterosinei C, (2R,3R)-pterosinei C, (2S,3R)-pterosinei C și (2R)-pterosinei B, cu valori IC50 de 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 și, respectiv, 29,6 μM. Ulterior, am testat potențialul inhibitor al derivaților de pterosin împotriva AChE. Toți compușii testați au prezentat activități semnificative de inhibare a AChE, cu valori IC50 în intervalul 2,55-110 μM, în comparație cu IC50 împotriva AChE a berberinei utilizate ca martor pozitiv, care a fost de 0,39 μM. Compușii de pterosină care au prezentat cea mai puternică activitate inhibitorie împotriva AChE au fost (2R)-pterosidul B, (2R,3R)-pterosidul C, (2S,3R)-pterosidul C, (2S,3R)-pterosidul C și (2R)-pterosidul B, cu valori IC50 de 2,55, 3,77, 9,17, 12,8 și, respectiv, 16,2 μM. În cele din urmă, am testat capacitățile inhibitorii ale derivaților de pterosin împotriva BChE. Toți compușii de pterosină testați au prezentat activitate inhibitoare împotriva BChE, cu valori IC50 care au variat de la 5,29 la 119 μM, cu excepția (3R)-pterosinei D care a fost inactivă la concentrațiile testate, în comparație cu IC50 a berberinei împotriva BChE, care a fost de 3,32 μM. Compușii pterosinici care au prezentat cea mai puternică activitate inhibitorie împotriva BChE au fost (2R,3R)-pterosidul C și pterosidul Z, cu valori IC50 de 5,29 și, respectiv, 5,31 μM.
Tabel 1 IC50 a derivaților de pterosină împotriva BACE1, AChE și BChE
Colectiv, majoritatea derivaților de pterosină testați au prezentat activități inhibitoare semnificative împotriva BACE1, AChE și BChE simultan. Prezența grupei suplimentare 2-hidroximetil-tetrahidro-piran-3,4,5-triol ca în derivații de pterosidă a crescut semnificativ activitățile inhibitorii împotriva enzimelor. Mai mult, prezența grupului hidroximetil suplimentar în poziția 2 a inelului indanonic al (2R)-pterosinei B, ca în (2S)-pterosina A, sau a grupului metil, ca în (3R)-pterosina D și pterosina Z, a redus activitățile inhibitoare împotriva enzimelor. În special, prezența grupării hidroximetil în poziția 5 a inelului indanonic ca în (2S)-pterosin P a scăzut activitatea inhibitorie față de BACE1.
Parametrii cinetici ai inhibiției enzimatice de către derivații de pterosină
În încercarea de a explica modul de inhibiție enzimatică a derivaților de pterosină, am efectuat o analiză cinetică a BACE1 și a colinesterazelor pentru inhibitori reprezentativi (tabelul 2, informații suplimentare 2). Un Ki (constantă de inhibiție) scăzut indică o legare mai strânsă a enzimei și un inhibitor mai eficient. În general, valorile Ki ale compușilor s-au corelat cu valorile IC50 respective. Inhibarea BACE1 de către compușii (2R,3R)-pteroside C, (3S)-pteroside D și (2R,3R)-pterosin C a fost de tip mixt, cu valori Ki de 12,6, 16,5 și 27.6 µM, respectiv, în timp ce inhibarea de către (2R)-pteroside B, (2S,3R)-pterosin C și (2R)-pterosin B a fost necompetitivă cu valori Ki de 23,1, 33,8 și, respectiv, 38,3 µM. Inhibarea AChE de către (2R)-pterosidul B, (2R,3R)-pterosidul C, (2R)-pterosidul B, (2S,3R)-pterosidul C și (3S)-pterosidul D a fost de tip mixt cu valori Ki de 4.89, 8,13, 12,1, 16,3 și, respectiv, 23,1 µM, în timp ce (2R,3R)-pterosina C a fost un inhibitor de tip necompetitiv cu o valoare Ki de 29,6 µM. Inhibarea BChE de către (2R,3R)-pterosin C, (2R,3R)-pterosid C, (3S)-pterosid D și (2R)-pterosid B a fost de tip mixt, cu valori Ki de 4,77, 9,62, 19.7 și, respectiv, 22,6 µM, în timp ce (2S,3R)-pterosina C și (2R)-pterosina B au fost inhibitori necompetitivi cu valori Ki de 29,9 și, respectiv, 53,5 µM. Astfel, aceste rezultate au sugerat că derivații specifici de pterosină ar putea fi inhibitori eficienți ai BACE1, AChE și BChE.
Tabelul 2 Cinetica enzimatică a derivaților de pterosină pe baza graficului Dixon și a graficului Lineweaver-Burk
Simulări de docking molecular pentru BACE1, AChE și BChE
Sunt disponibile mai multe structuri cristaline pentru BACE1 și colinesteraze. Am selectat PDB umane pe baza structurilor de tip sălbatic, a liganzilor cocristalizați și a rezoluțiilor structurilor. Structurile cristaline cu raze X ale BACE1 complexate cu QUD (cod PDB: 2WJO, rezoluție: 2,5 Å)33, ale AChE complexate cu E2020 (cod PDB: 4EY7, rezoluție: 2,35 Å)34 și ale BChE complexate cu 3F9 (cod PDB: 4TPK, rezoluție: 2,70 Å)35 au fost selectate pentru andocare. Inițial, QUD, E2020 și 3F9 au fost extrase din structurile cristaline și redockate în situsurile active ale BACE1, AChE și, respectiv, BChE. Ulterior, (2R,3R)-pterozidul C, (3S)-pterozidul D, (2R,3R)-pterozidul C, (2R)-pterozidul B, (2S,3R)-pterozidul C și (2R)-pterozidul B cu mecanismul cunoscut de inhibiție împotriva BACE1, AChE și BChE au fost andocați pentru a determina SAR-ul acestora. Rezultatele docking-ului sunt rezumate în tabelul 3. SAR-ul inhibitorilor BACE1, AChE și BChE de tip mixt și necompetitiv selectați a permis evaluarea unei noi inhibiții a enzimelor de către pterosine, dirijate de locul de legare.
Tabelul 3 Scorurile de afinitate de andocare și posibila formare a legăturilor H la situsurile active corespunzătoare ale BACE1, AChE și BChE de către derivații de pterosină împreună cu inhibitorii raportați
Modul nostru de andocare al E2020 a fost în concordanță cu modul de legare determinat experimental raportat anterior cu AChE umană recombinantă (rhAChE) (Informații suplimentare 3)34. Abaterea medie pătratică (RMSD) între conformația cristalină și conformația de andocare a E2020 a fost de 0,54 Å, ceea ce sugerează fiabilitatea configurației noastre de andocare în reproducerea modului de legare experimental. În plus, modul de andocare a E202020 a condus la o interacțiune similară cu cea a rhAChE-E2020. În studiul nostru, moleculele de apă au fost eliminate din structura cristalină în timpul docking-ului; prin urmare, interacțiunile mediate de apă nu au fost analizate în studiul de față. În mod similar, modurile de andocare ale QUD și 3F9 au fost în concordanță cu datele experimentale disponibile pentru BACE1 33 și, respectiv, BChE35 (Informații suplimentare 3). RMSD între conformațiile cristaline și cele andocate ale QUD și 3F9 au fost de 0,46 și, respectiv, 0,60 Å. Mai mult, situsurile de legare ale inhibitorilor de pterosină au fost în concordanță cu un studiu de andocare anterior care a implicat BACE1, AChE și BChE38. Cu toate acestea, studiul a utilizat Tetronarce californica AChE (cod PDB: 1ACJ), care conține numere de reziduuri ușor diferite față de AChE umană din cauza variațiilor în secvențele lor.
BACE1 docking
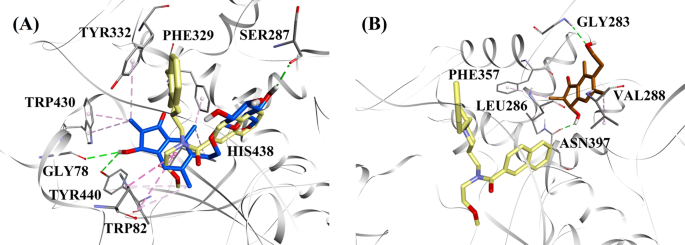
Pe baza tipului de inhibiție și a activității, au fost selectate (2R,3R)-pterosida C și (2R)-pterosida B pentru a demonstra modurile de docking ale inhibitorilor BACE1 de tip mixt și, respectiv, necompetitiv. Figura 3a, b prezintă modelele de andocare a (2R,3R)-pterosidei C și, respectiv, a (2R)-pterosidei B. Interacțiunile compușilor andocați în interiorul situsului activ al BACE1 sunt prezentate în Fig. 4.
Fig. 3: Modele de andocare moleculară pentru inhibitorii BACE1 de tip mixt și cei necompetitivi.
Modeluri de andocare moleculară pentru a inhibitorul BACE1 de tip mixt (2R,3R)-pterosid C (culoare albastră) și b inhibitorul BACE1 necompetitiv (2R)-pterosid B (culoare maro). Pozițiile andocate sunt suprapuse peste structura cristalină cu raze X a QUD (culoare galbenă) (cod PDB: 2WJO). BACE1, reziduurile situsului activ și compușii sunt reprezentați prin modele de tip panglică, linie și, respectiv, băț. Culorile liniilor punctate explică tipurile de diverse interacțiuni: interacțiuni de legătură de hidrogen (verde), interacțiuni hidrofobe (roz) și interacțiuni π-sigma (violet). BACE1 β-site-ul enzimei de clivare a proteinei precursoare amiloide 1
Fig. 4: Diagrama de interacțiune cu ligandul a inhibitorilor BACE1 în situsul activ.
Diagrama de interacțiune a ligandului a QUD, b (2R,3R)-pteroside C, c (3S)-pteroside D, d (2R,3R)-pterosin C, e (2R)-pteroside B, f (2S,3R)-pterosin C, și g (2R)-pterosin B în situsul activ al BACE1. Culorile liniilor punctate explică tipurile diferitelor interacțiuni: interacțiuni de legătură de hidrogen (verde), interacțiuni hidrofobe (roz) și interacțiuni π-sigma (violet). BACE1 β-site-ul enzimei de clivare a proteinei precursoare amiloide 1
Postura andocată a QUD a prezentat o energie de legătură (B.E.) de -7,59 kcal/mol. După cum se arată în Fig. 4a, grupul NH2 de pe inelul de quinazolină al ligandului a prezentat două legături de hidrogen cu grupurile CO ale Asp32 și Asp228 la distanțe de 1,86 și, respectiv, 2,17 Å. O a treia legătură de hidrogen a fost observată între celălalt grup NH al ligandului și grupul CO al Gly230 la o distanță de 2,16 Å. Leu30, Tyr71, Phe108 și Val332 au mediat interacțiunile hidrofobe. În figura 4b-d sunt afișate ipostazele docked ale (2R,3R)-pterosidei C, (3S)-pterosidei D și, respectiv, ale (2R,3R)-pterosinei C (inhibitori BACE1 de tip mixt). Aceștia au fost poziționați în buzunarul de legare delimitat de Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 și Thr231. În ceea ce privește nivelurile lor de activitate, (2R,3R)-pterosidul C (IC50 = 9,74 µM), (3S)-pterosidul D (IC50 = 10,7 µM) și (2R,3R)-pterosina C (IC50 = 26,2 µM) au prezentat un B.E. de -7,27, -6,93 și, respectiv, -4,84 kcal/mol. (2R,3R)-Pterosidul C a prezentat o potență mai mare decât (2R,3R)-pterosina C datorită existenței unei grupări suplimentare de 2-hidroximetil-tetrahidro-piran-3,4,5-triol, care a format trei legături de hidrogen (Fig. 4b). Grupul OH al hidroximetilului și grupul 3-OH al inelului tetrahidro-piran-triol au prezentat două legături de hidrogen cu grupul CO al Asp228 la distanțe de 2,63 și, respectiv, 2,12 Å. În plus, grupul 3-OH a demonstrat o legătură de hidrogen cu Thr231 la o distanță de 1,89 Å. În cazul (3S)-pterosidei D (Fig. 4c), prezența grupului 2,2-dimetil la inelul indanonic a modificat ușor interacțiunile de legare în comparație cu (2R,3R)-pterosida C. Grupul 3-OH al inelului tetrahidro-piran-triol a prezentat doar o singură legătură de hidrogen cu Asp228 la o distanță de 1,93 Å. Cu toate acestea, grupul 3-OH al inelului de indanonă a prezentat o legătură de hidrogen suplimentară cu Ile126 la o distanță de 2,34 Å. Aceste interacțiuni au redus ușor activitatea (3S)-pterosidei D în comparație cu (2R,3R)-pterosidei C. Interacțiunile de andocare ale (2R,3R)-pterosidei C afișate în Fig. 4d arată că gruparea 3-OH a inelului de indanonă este legată de Ser36 și Asn37 prin intermediul a două legături de hidrogen la distanțe de 1,83 și, respectiv, 2,07 Å. Ala39, Val69, Trp76, Ile118 și Arg128 au fost implicate în interacțiuni hidrofobe, în timp ce Val69 a prezentat o interacțiune π-sigma.
Posturile docked ale (2R)-pterosidei B, (2S,3R)-pterosinei C și (2R)-pterosinei B (inhibitori necompetitivi ai BACE1) sunt prezentate în Fig. 4e-g, respectiv. Aceștia au fost andocați în cavitatea delimitată de Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 și Ile126. În conformitate cu valorile lor de activitate, (2R)-pterosidul B (IC50 = 18,0 µM), (2S,3R)-pterosina C (IC50 = 23,1 µM) și (2R)-pterosina B (IC50 = 29,6 µM) au prezentat un B.E. de -6,16, -5,07 și, respectiv, -4,64 kcal/mol. După cum se arată în Fig. 4e, (2R)-pterosidul B a demonstrat o activitate mai mare decât (2S,3R)-pterosina C și (2R)-pterosina B datorită prezenței unei grupări suplimentare de 2-hidroximetil-tetrahidro-piran-3,4,5-triol, care a prezentat patru interacțiuni de legături de hidrogen. Au fost observate două legături de hidrogen între grupul 4-OH al inelului tetrahidro-piran-triol și grupurile NH și CO ale Asn37 la distanțe de 2,48 și, respectiv, 2,19 Å. În plus, grupurile 3 și 5-OH au prezentat două legături de hidrogen suplimentare cu Ile126 și Trp76 la distanțe de 2,19 și, respectiv, 2,46 Å. (2S,3R)-Pterosina C a prezentat o activitate ușor mai bună decât (2R)-pterosina B datorită prezenței unei grupări OH suplimentare în poziția 3 a inelului de indanonă, care a format o legătură de hidrogen cu Lys107 la o distanță de 2,12 Å (Fig. 4f). Celelalte interacțiuni au fost similare cu cele ale (2R)-pterosinei B. După cum se arată în Fig. 4g, (2R)-pterosina B a prezentat interacțiuni hidrofobe cu Val69, Tyr76 și Phe108.
AChE docking
(2R)-Pteroside B și (2R,3R)-pterosin C au fost selectate ca reprezentanți pentru a demonstra modurile docked ale inhibitorilor AChE de tip mixt și, respectiv, necompetitiv, datorită activităților și tipului lor de inhibiție AChE. Figura 5a, b ilustrează modelele de andocare ale (2R)-pterosidei B și, respectiv, ale (2R,3R)-pterosinei C. Interacțiunile compușilor andocați în interiorul situsului activ al AChE sunt prezentate în Fig. 6.
Fig. 5: Modele de andocare moleculară pentru inhibitorii AChE de tip mixt și necompetitiv.
Modeluri de andocare moleculară pentru a inhibitorul AChE de tip mixt (2R)-pteroside B (culoare albastră) și b inhibitorul AChE necompetitiv (2R,3R)-pterosin C (culoare maro). Pozițiile andocate sunt suprapuse peste structura cristalină cu raze X a E2020 (culoare galbenă) (cod PDB: 4EY7). AChE, reziduurile situsului activ și compușii sunt reprezentați prin modele de tip panglică, linie și, respectiv, băț. Culorile liniilor punctate explică tipurile de diverse interacțiuni: interacțiuni de legătură de hidrogen (verde) și interacțiuni hidrofobe (roz). AChE acetilcolinesterază
Fig. 6: Diagrama de interacțiune cu ligandul a inhibitorilor AChE în situsul activ.
Diagrama de interacțiune a ligandului a E2020, b (2R)-pteroside B, c (2R,3R)-pteroside C, d (2S,3R)-pterosin C, e (2R)-pterosin B, f (3S)-pteroside D și g (2R,3R)-pterosin C în situsul activ al AChE. Culorile liniilor punctate explică tipurile diferitelor interacțiuni: interacțiuni de legătură de hidrogen (verde), interacțiuni hidrofobe (roz) și interacțiuni π-sigma (violet). AChE acetilcolinesterază
Postura andocată a lui E2020 a demonstrat un B.E. de -10,28 kcal/mol. După cum este ilustrat în Fig. 6a, gruparea CO a inelului indanonic a format o legătură de hidrogen cu gruparea NH a Phe295 la o distanță de 1,70 Å. Trp286 și Tyr341 au fost implicate în interacțiuni π-sigma, în timp ce Trp86, Trp286, Tyr337, Phe338 și Tyr341 au mediat interacțiuni hidrofobe. Figura 6b-f demonstrează posturile docked ale (2R)-pterosidei B, (2R,3R)-pterosidei C, (2S,3R)-pterosinei C, (2R)-pterosinei B și, respectiv, (3S)-pterosidei D (inhibitori AChE de tip mixt). Aceștia au fost găzduiți în situsul activ înconjurat de Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 și Tyr341. În concordanță cu valorile lor de activitate, (2R)-pterozidul B (IC50 = 2,55 µM), (2R,3R)-pterozidul C (IC50 = 3,77 µM), (2S,3R)-pterozidul C (IC50 = 12.8 µM), (2R)-pterosin B (IC50 = 16,2 µM) și (3S)-pteroside D (IC50 = 27,4 µM) au prezentat un B.E. de -7,90, -7,49, -6,03, -5,76 și, respectiv, -4,91 kcal/mol. (2R)-Pterosida B a demonstrat o potență mai mare decât (2S,3R)-pterosina C și (2R)-pterosina B datorită prezenței unei grupări suplimentare de 2-hidroximetil-tetrahidro-piran-3,4,5-triol, care a stabilit trei interacțiuni de legături de hidrogen (Fig. 6b). Grupul OH al hidroximetilului și grupul 3-OH al inelului tetrahidro-piran-triol au prezentat legături de hidrogen cu grupul CO al Asn87 la distanțe de 2,23 și, respectiv, 2,28 Å. Mai mult, grupul 4-OH a format o legătură de hidrogen cu grupul CO al Trp86 la o distanță de 2,13 Å. În cazul (2R,3R)-pterosidei C (Fig. 6c), grupul metil din poziția 2 a inelului de indanonă nu a prezentat o interacțiune hidrofobă cu Tyr341 și, prin urmare, a prezentat o activitate comparativ mai scăzută decât (2R)-pterosida B. Cu toate acestea, Tyr341 a menținut interacțiunea hidrofobă cu cealaltă parte a inelului de indanonă, așa cum se arată în cazul (2R)-pterosidei B. Activitatea mai mare a (2S,3R)-pterosinei C decât a (2R)-pterosinei B a fost atribuită existenței unei grupări OH suplimentare în poziția-3 a inelului de indanonă, care a format o legătură de hidrogen cu Phe295 la o distanță de 1,82 Å (Fig. 6d). Celelalte interacțiuni au fost comparabile cu interacțiunile (2R)-pterosin B. După cum se arată în Fig. 6e, grupul OH al grupului hidroxietil din poziția 6 a inelului indanonic a format o legătură de hidrogen cu Tyr124 la o distanță de 2,48 Å. Tyr124, Trp286, Tyr337, Phe338 și Tyr341 au contribuit la interacțiunile hidrofobe. În comparație cu (2R)-pterozidul B și (2R,3R)-pterozidul C, (3S)-pterozidul D a prezentat interacțiuni de legare diferite din cauza prezenței grupului 2,2-dimetil la inelul de indanonă (Fig. 6f). Grupul 2,2-dimetil a contribuit în mod semnificativ la poziția de andocare distinctă a (3S)-pterosidei D. Grupul 5-OH al inelului tetrahidro-piran-triol a format o legătură de hidrogen cu Asp74 la o distanță de 1,96 Å. Grupul CO al inelului de indanonă a prezentat o legătură de hidrogen cu Tyr72 la o distanță de 2,97 Å. Aceste interacțiuni au explicat activitatea scăzută a (3S)-pterosidei D.
Postura andocată a (2R,3R)-pterosinei C (inhibitor necompetitiv al AChE) este prezentată în Fig. 6g. (2R,3R)-Pterosina C (IC50 = 23,2 µM) a demonstrat un B.E. de -5,01 kcal/mol. Buzunarul de legare al (2R,3R)-pterosinei C cuprindea Tyr72, Trp286, Ser293, Phe297 și Tyr341, cu două interacțiuni prin legături de hidrogen. Una dintre legăturile de hidrogen a fost formată între grupul 3-OH al inelului indanonic și grupul CO al Ser293 la o distanță de 2,03 Å. Cea de-a doua legătură de hidrogen a fost observată între grupul OH al grupului hidroxietil prezent în poziția 6 a inelului indanonic și Tyr72 la o distanță de 1,94 Å. Reziduuri precum Tyr72, Trp286, Phe297 și Tyr341 au participat la interacțiuni hidrofobe.
Considerând nivelurile de activitate și tipul de inhibiție a BChE, (2R,3R)-pteroside C și (2S,3R)-pterosin C au fost selectate pentru a demonstra modurile de andocare a inhibitorilor BChE de tip mixt și, respectiv, necompetitiv. Figura 7a, b ilustrează modelele de andocare ale (2R,3R)-pterosidei C și, respectiv, ale (2S,3R)-pterosinei C. Interacțiunile compușilor andocați în interiorul situsului activ BChE sunt prezentate în Fig. 8.
Fig. 7: Modele de andocare moleculară pentru inhibitorii BChE de tip mixt și necompetitivi.
Modele de docking molecular pentru a inhibitorul BChE de tip mixt (2R,3R)-pterosid C (culoare albastră) și b inhibitorul BChE necompetitiv (2S,3R)-pterosin C (culoare maro). Pozițiile andocate sunt suprapuse peste structura cristalină cu raze X a 3F9 (culoare galbenă) (cod PDB: 4TPK). BChE, reziduurile situsului activ și compușii sunt reprezentați prin modele de tip panglică, linie și, respectiv, băț. Culorile liniilor punctate explică tipurile de diverse interacțiuni: interacțiuni de legătură de hidrogen (verde) și interacțiuni hidrofobe (roz). BChE butirilcolinesterază
Fig. 8: Diagrama de interacțiune cu ligandul a inhibitorilor BChE în situsul activ.
Diagrama de interacțiune a ligandului a 3F9, b (2R,3R)-pterosid C, c (3S)-pterosid D, d (2R,3R)-pterosin C, e (2R)-pterosid B, f (2S,3R)-pterosin C și g (2R)-pterosin B în situsul activ al BChE. Culorile liniilor punctate explică tipurile diferitelor interacțiuni: interacțiuni de legătură de hidrogen (verde), interacțiuni hidrofobe (roz) și interacțiuni π-anion (auriu). BChE butirilcolinesterază
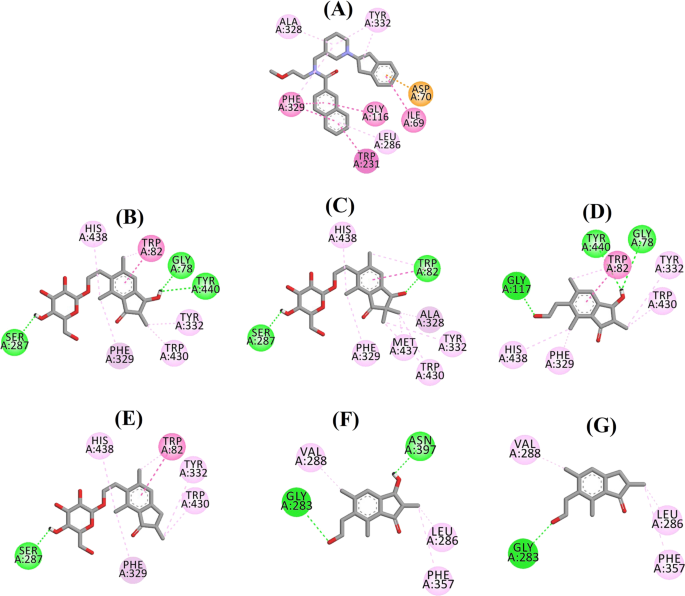
Postura andocată a lui 3F9 a arătat un B.E. de -8,49 kcal/mol. După cum se arată în Fig. 8a, interacțiunile hidrofobe au fost în principal responsabile pentru legarea ligandului. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 și Tyr332 au reprezentat interacțiunile hidrofobe, în timp ce Asp70 a demonstrat o interacțiune π-anion. Figura 8b-e ilustrează ipostazele docked ale (2R,3R)-pterosidei C, (3S)-pterosidei D, (2R,3R)-pterosinei C și, respectiv, (2R)-pterosidei B (inhibitori BChE de tip mixt). Buzunarul lor de legare a fost compus din Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 și Tyr440. În conformitate cu nivelurile lor de activitate, (2R,3R)-pterosidul C (IC50 = 5,29 µM), (3S)-pterosidul D (IC50 = 19,3 µM), (2R,3R)-pterosina C (IC50 = 20,3 µM) și (2R)-pterosidul B (IC50 = 62,0 µM) au demonstrat un B.E. de -7,23, -6,59, -6,52 și, respectiv, -4,38 kcal/mol. După cum se arată în Fig. 8b, grupul 3-OH al inelului indanonic al (2R,3R)-pterosidei C a prezentat două legături de hidrogen cu Gly78 și Tyr440 la distanțe de 2,87 și, respectiv, 2,89 Å. În cazul (3S)-pterosidei D, grupul 2,2-dimetil de pe inelul indanonic a afectat interacțiunile de legătură ale grupului 3-OH (Fig. 8c). Grupul 3-OH a format doar o singură legătură de hidrogen cu Trp82 la o distanță de 2,94 Å, ceea ce a dus la o activitate comparativ mai scăzută a (3S)-pterosidei D decât a (2R,3R)-pterosidei C. După cum se arată în Fig. 8d, (2R,3R)-pterosina C nu a reușit să producă o legătură de hidrogen cu Ser287 din cauza absenței unui grup 2-hidroximetil-tetrahidro-piran-3,4,5-triol. În consecință, a dat o activitate mai scăzută decât (2R,3R)-pterosidul C. S-a constatat că lipsa grupului OH în poziția 3 a inelului indanonic este responsabilă pentru activitatea foarte scăzută a (2R)-pterosidului B (Fig. 8e), care nu a reușit să formeze legături de hidrogen cu Gly78 și Tyr440 similar cu (2R,3R)-pterosidul C.
Posturile docked ale (2S,3R)-pterosinei C și (2R)-pterosinei B (inhibitori necompetitivi ai BChE) sunt prezentate în Fig. 8f, respectiv g. Aceste poziții andocate au fost conținute în cavitatea delimitată de Gly283, Leu286, Val288, Phe357 și Asn397. În ceea ce privește nivelurile lor de activitate, (2S,3R)-pterosina C (IC50 = 44,3 µM) și (2R)-pterosina B (IC50 = 48,1 µM) au prezentat un B.E. de -5,40 și, respectiv, -5,06 kcal/mol. (2S,3 R)-Pterosina C a fost mai puternică decât (2R)-pterosina B din cauza prezenței unei grupări OH suplimentare în poziția 3 a inelului indanonic, care a format o legătură de hidrogen cu gruparea CO a Asn397 la o distanță de 2,05 Å (Fig. 8f). Alte interacțiuni s-au dovedit a fi similare cu (2R)-pterosina B. După cum se arată în Fig. 8g, grupul OH din grupul hidroxietil de la poziția 6 a inelului indanonic a format o legătură de hidrogen cu Gly283 la o distanță de 2,40 Å. Reziduurile Leu286, Val288 și Phe357 au participat la interacțiuni hidrofobe.
Inhibitorii de tip mixt se leagă atât la enzima liberă, cât și la complexul enzimă-substrat, ceea ce indică faptul că acești compuși se pot lega la situsul catalitic al fiecărei enzime corespunzătoare. Inhibitorii necompetitivi se leagă la situsul alosteric al enzimei libere sau al complexului enzimă-substrat. Un studiu recent a sugerat că inhibitorii competitivi, de tip mixt și necompetitivi ocupă locuri diferite în buzunarele de legare ale BACE1, AChE și BChE38. În timpul docking-ului pentru evaluarea mecanismului inhibitor al derivaților de pterosină, situsurile de legare ale compușilor au fost definite în funcție de tipul lor de inhibiție. Rezultatele docking-ului au indicat că situsurile de legare ale inhibitorilor de tip mixt și necompetitiv pentru BACE1, AChE și BChE se suprapun parțial unul peste celălalt la fiecare situs activ corespunzător și au fost în concordanță cu un raport anterior38.
Permeabilitatea BBB
PAMPA-BBB, un test de permeabilitate a membranei artificiale in vitro pentru BBB, este unul dintre cele mai fiabile instrumente de screening fizico-chimic în etapa inițială de descoperire a medicamentelor care vizează SNC40. Sistemul PAMPA-BBB modelează difuzia pasivă transcelulară a substanțelor chimice prin BBB și măsoară mecanismele de transport strict pasiv prin intermediul unei membrane lipidice artificiale asupra permeabilității efective (Pe, cm/s). Pe baza modelului stabilit pentru predicția permeabilității BBB, compușii au fost clasificați în (i) „CNS+” (predicție ridicată a permeabilității BBB); Pe (10-6 cm/s) > 4,00, (ii) „CNS-” (predicție scăzută a permeabilității BBB); Pe (10-6 cm/s) < 2,00 și (iii) „CNS+/-” (permeabilitate BBB incertă); Pe (10-6 cm/s) de la 4,00 la 2,00. În consecință, (2R)-pterosina B, (2S)-pterosina P și (2S)-pterosina A au prezentat o permeabilitate BBB ridicată, cu valori Pe de 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s și, respectiv, 6,26 × 10-6 cm/s (tabelul 4, informații suplimentare 4). Valoarea Pe a (2R)-pterosinei B a fost de 1,7 ori mai mare decât cea a medicamentului pentru SNC verapamil (Pe = 34,6 × 10-6 cm/s), care a fost utilizat pentru controlul pozitiv. (2S,3R)-Pterosina C și (2R,3R)-pterosina C au prezentat o permeabilitate incertă în BBB cu valori Pe de 2,34 și, respectiv, 1,98. (2R,3R)-Pterosidul C, (3S)-pterosidul D și (2R)-pterosidul B, care au prezentat cele mai puternice activități de inhibare a BACE1 și a colinesterazei dintre derivații de pterosină testați, au prezentat o permeabilitate BBB foarte scăzută. Existența grupului 2-hidroximetil-tetrahidro-piran-3,4,5-triol ca în cazul derivaților de pterosidă a dus la o scădere remarcabilă a permeabilității BBB. Comparativ cu (2R)-pterosina B, prezența suplimentară a grupului OH în poziția 3 a inelului indanonic ca în pterosina C, a grupului hidroximetil în poziția 2 a inelului indanonic ca în (2S)-pterosina A sau a grupului hidroximetil în poziția 5 a inelului indanonic ca în (2S)-pterosina P a redus, de asemenea, în mod semnificativ permeabilitatea BBB. Având în vedere o permeabilitate BBB excepțional de ridicată și inhibarea semnificativă a BACE1, AChE și BChE, (2R)-pterosina B poate avea potențialul de a prezenta o puternică activitate anti-AD.
Tabel 4 Permeabilitatea PAMPA-BBB a derivaților de pterosină
Efectele (2R)-pterosinei B și (2R,3R)-pterosidei C asupra secreției de peptide Aβ de către celulele neuronale
Pentru a investiga funcția (2R)-pterosinei B și (2R,3R)-pteroside C în diminuarea excreției de Aβ de către celulele neuronale, am utilizat o linie celulară de neuroblastom murin care supraexprimă în mod stabil APPswe uman. Linia celulară este un model celular de DA caracterizat prin secreția excesivă de Aβ40 și Aβ42. Oligomerii amiloizi toxici sunt formați din cele două izoforme ale peptidei Aβ cu lungimi diferite. Aβ40 este cea mai abundentă izoformă Aβ în creier41, în timp ce Aβ42 crește semnificativ în cazul anumitor forme de DA42. Testul ELISA sandwich pentru Aβ40 a arătat că (2R)-pterosina B a redus semnificativ cantitatea de peptidă Aβ40 secretată din celulele de neuroblastom în mediu cu până la 50 % la 500 μM (P < 0,01) (Fig. 9a) (Fig. 9a). În mod similar, secreția de peptidă Aβ42 de către celulele de neuroblastom a scăzut semnificativ în prezența a 500 μM de (2R,3R)-pterosidă C (P < 0,05) (Fig. 9b). În concluzie, (2R)-pterosina B și (2R,3R)-pterosida C au diminuat semnificativ secreția de peptide Aβ de către celulele de neuroblastom la o concentrație de 500 μM.
Fig. 9: Efectele (2R)-pterosinei B și (2R,3R)-pterosidei C asupra secreției celulare de peptide Aβ.
a Efectul (2R)-pterosinei B asupra secreției de peptide Aβ. Celulele de neuroblastom care secretă APPswe au fost tratate cu 12, 60, 250 sau 500 µM de (2R)-pterosin B timp de 24 h, iar mediile condiționate au fost colectate în prezența unui inhibitor de protează. β-SI (10 µM) a fost utilizat ca martor pozitiv. Controlul negativ a inclus celule cultivate în absența compușilor de testare. Analiza cantitativă a Aβ40 și Aβ42 secretate în mediile condiționate a fost efectuată cu ajutorul testului ELISA sandwich. Sunt prezentate mediile ± SD din trei experimente independente. Peptidele Aβ40 secretate au scăzut semnificativ în prezența a 500 µM (2R)-pterosin B. CON control negativ, β-SI β-inhibitor III de secretază, Aβ β-amiloid. *P < 0,05, **P < 0,01. b Efectul (2R,3R)-pterosidei C asupra secreției de peptide Aβ. Celulele de neuroblastom care secretă APPswe au fost tratate cu 12, 60, 125, 250 sau 500 µM de (2R,3R)-pterosid C timp de 24 h, iar mediile condiționate au fost colectate în prezența unui inhibitor de protează. β-SI (10 µM) a fost utilizat ca martor pozitiv. Controlul negativ a inclus celule cultivate în absența compușilor de testare. Analiza cantitativă a Aβ40 și Aβ42 secretate în mediile condiționate a fost efectuată cu ajutorul testului ELISA sandwich. Sunt prezentate mediile ± SD din trei experimente independente. Peptida Aβ42 secretată a scăzut semnificativ în prezența a 500 µM (2R,3R)-pterosid C. CON control negativ, β-SI β-inhibitor III de secretază, Aβ β-amiloid. *P < 0.05
Citotoxicitatea derivaților de pterosină pe baza testului MTT43
În general, derivații de pterosină au prezentat o citotoxicitate neglijabilă față de diverse linii celulare normale și canceroase, cum ar fi neuroblastomul uman SH-SY5Y, celulele gliale de șobolan C6, fibroblastele embrionare de șoarece NIH3T3 și melanomul de șoarece B16F10, cu valori LD50 peste 0.5 mM (Informații suplimentare 5). În special, (2R)-pterosina B, (3R)-pterosina D, (2S)-pterosina P, (2S)-pterosidul A, (2R)-pterosidul B și (2R,3R)-pterosidul C nu au prezentat citotoxicitate împotriva liniilor celulare testate cu valori LD50 de peste 5 mM. În mod ciudat, mai multe pterosine au prezentat efecte antiproliferative relative împotriva celulelor neuronale SH-SY5Y în comparație cu celulele gliale C6 de origine mezenchimală. Rezultatele prezente au fost în concordanță cu un raport anterior care a sugerat că derivații de pterosină nu sunt toxici pentru oameni26.