

Contribuția unei punți de sare la stabilitatea generală la starea pliată a unei proteine poate fi evaluată prin date termodinamice colectate din studii de mutageneză și tehnici de rezonanță magnetică nucleară. Utilizând o proteină mutantă de tip pseudo-wild, mutată în mod specific pentru a preveni precipitarea la pH ridicat, contribuția punții saline la energia liberă globală a stării de pliere a proteinei poate fi determinată prin efectuarea unei mutații punctiforme, modificând și, în consecință, rupând puntea salină. De exemplu, s-a identificat existența unei punți saline în lizozima T4 între acidul aspartic (Asp) de la reziduul 70 și o histidină (His) de la reziduul 31 (figura 3). S-a efectuat o mutageneză dirijată la fața locului cu asparagină (Asn) (figura 4), obținându-se trei noi mutanți: Asp70Asn His31 (Mutant 1), Asp70 His31Asn (Mutant 2), și Asp70Asn His31Asn (Mutant dublu).

Odată ce au fost stabiliți mutanții, pot fi folosite două metode pentru a calcula energia liberă asociată unei punți saline. O metodă implică observarea temperaturii de topire a proteinei de tip sălbatic față de cea a celor trei mutanți. Denaturarea poate fi monitorizată printr-o modificare a dicroismului circular. O reducere a temperaturii de topire indică o reducere a stabilității. Aceasta este cuantificată printr-o metodă descrisă de Becktel și Schellman, în care diferența de energie liberă dintre cele două este calculată prin ΔTΔS. Există unele probleme legate de acest calcul și poate fi utilizat numai cu date foarte precise. În exemplul lizozimei T4, ΔS de tip pseudo-wild a fost raportată anterior la pH 5,5, astfel încât diferența de temperatură mediană de 11 °C la acest pH înmulțită cu ΔS raportată de 360 cal/(mol-K) (1,5 kJ/(mol-K)) produce o schimbare de energie liberă de aproximativ -4 kcal/mol (-17 kJ/mol). Această valoare corespunde cantității de energie liberă contribuită la stabilitatea proteinei de către puntea salină.
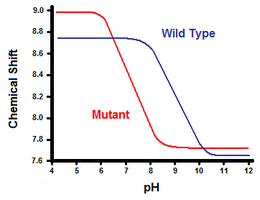
A doua metodă utilizează spectroscopia de rezonanță magnetică nucleară pentru a calcula energia liberă a punții saline. Se efectuează o titrare, înregistrând în același timp deplasarea chimică corespunzătoare protonilor carbonului adiacent grupului carboxilat sau amoniu. Punctul median al curbei de titrare corespunde pKa, sau pH-ului la care raportul dintre moleculele protonate și cele deprotonate este de 1:1. Continuând cu exemplul lizozimei T4, se obține o curbă de titrare prin observarea unei deplasări a protonului C2 al histidinei 31 (figura 5). Figura 5 arată deplasarea curbei de titrare între tipul sălbatic și mutantul în care Asp70 este Asn. Puntea salină formată este între Asp70 deprotonat și His31 protonat. Această interacțiune determină deplasarea observată în pKa lui His31. În proteina de tip sălbatic desfășurată, în care puntea salină este absentă, His31 are un pKa de 6,8 în tampoane H20 de forță ionică moderată. Figura 5 arată un pKa al tipului sălbatic de 9,05. Această diferență de pKa este susținută de interacțiunea lui His31 cu Asp70. Pentru a menține puntea salină, His31 va încerca să își păstreze protonul cât mai mult timp posibil. Atunci când puntea salină este întreruptă, ca în cazul mutantului D70N, pKa revine la o valoare de 6,9, mult mai apropiată de cea a His31 în stare desfășurată.
Diferența în pKa poate fi cuantificată pentru a reflecta contribuția punții saline la energia liberă. Folosind energia liberă Gibbs:ΔG = -RT ln(Keq), unde R este constanta universală a gazelor, T este temperatura în kelvins, iar Keq este constanta de echilibru a unei reacții în echilibru. Deprotonarea lui His31 este o reacție acidă de echilibru cu o Keq specială, cunoscută sub numele de constanta de disociere a acizilor, Ka: His31-H+ ⇌ His31 + H+. PKa este apoi legată de Ka prin următoarea relație: pKa = -log(Ka). Calculul diferenței de energie liberă a mutantului și a tipului sălbatic se poate face acum folosind ecuația energiei libere, definiția pKa, valorile pKa observate și relația dintre logaritmii naturali și logaritmi. În exemplul lizozimei T4, această abordare a dus la o contribuție calculată de aproximativ 3 kcal/mol la energia liberă globală. O abordare similară poate fi adoptată cu celălalt participant la puntea salină, cum ar fi Asp70 în exemplul lizozimei T4, prin monitorizarea deplasării sale în pKa după mutația lui His31.
Un cuvânt de precauție la alegerea experimentului adecvat implică localizarea punții saline în cadrul proteinei. Mediul înconjurător joacă un rol important în interacțiune. La concentrații ionice ridicate, puntea salină poate fi complet mascată, deoarece este implicată o interacțiune electrostatică. Puntea salină His31-Asp70 din lizozima T4 a fost îngropată în interiorul proteinei. Entropia joacă un rol mai important în cazul punților saline de suprafață, unde reziduurile care în mod normal au capacitatea de a se mișca sunt constrânse de interacțiunea lor electrostatică și de legătura de hidrogen. S-a demonstrat că acest lucru scade entropia suficient de mult pentru a șterge aproape complet contribuția interacțiunii. Punctele de sare de suprafață pot fi studiate în mod similar cu cele ale punților de sare îngropate, utilizând cicluri cu dublu mutant și titrări RMN. Deși există cazuri în care punțile de sare îngropate contribuie la stabilitate, ca orice altceva, există și excepții, iar punțile de sare îngropate pot prezenta un efect destabilizator. De asemenea, punțile de sare de suprafață, în anumite condiții, pot prezenta un efect stabilizator. Efectul stabilizator sau destabilizator trebuie evaluat de la caz la caz și se pot face puține afirmații generale.
.