
Apasningens miljømæssige kontekst
Vi har mange gange før talt om, hvordan arter udvikler sig som reaktion på en form for miljømæssigt pres, der begunstiger (eller ikke begunstiger) visse træk inden for den pågældende art. Med tiden fører dette til ændringer i hyppigheden af arternes træk og ændrer den samlede gennemsnitlige fænotype for den pågældende art (nogle gange langsomt, andre gange hurtigt).
Mens vi normalt taler om miljøet i form af abiotiske forhold som temperatur eller klima, er biotiske faktorer lige så vigtige: det vil sige de dele af miljøet, som også selv er levende. Derfor kan ændringer hos én art få store konsekvenser for andre arter, der er knyttet til økosystemet. Udviklingen af en art er således uløseligt forbundet med udviklingen af andre relevante arter i økosystemet: ofte kæmper disse forbundne udviklingsveje mod hinanden, når de hver især ændrer sig. Lad os tage et kig på et par forskellige eksempler på, hvordan udviklingen af en art kan påvirke udviklingen af en anden art.
Røver-bytte-koevolution
En af de mest åbenlyse måder, hvorpå udviklingen af to forskellige arter kan interagere, er i forholdet mellem rovdyr og bytte. Naturligvis udvikler byttearter sig for at kunne forsvare sig mod rovdyr på forskellige måder, f.eks. ved hjælp af crypsis (f.eks. camouflage), giftighed eller adfærdsændringer (f.eks. natteaktivitet eller gruppedrift). Omvendt vil rovdyrene udvikle nye og forbedrede metoder til at opdage og jage byttet, f.eks. forbedrede sanser, gift og snigeri (f.eks. gennem blødpolstrede fødder).
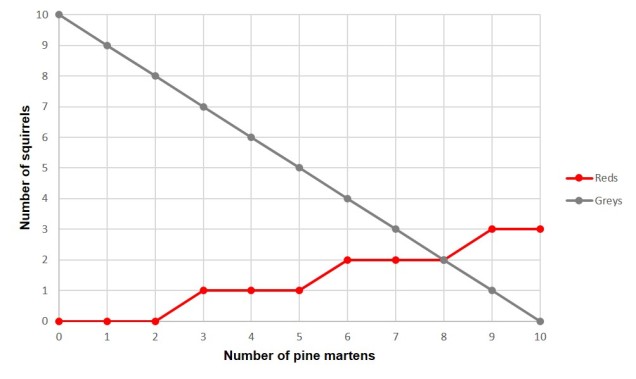
Der er millioner af mulige eksempler på samudvikling mellem rovdyr og bytte, som kunne bruges som eksempler her, baseret på den kontinuerlige trang for den ene art til at få overtaget over den anden. Men et af dem, som jeg kommer til at tænke på, er et dyr, som jeg lærte om under en ferie i Skandinavien, nemlig skovmåren, og hvordan den påvirker egern.

Fyrre-måren er en art i mustelid-familien sammen med oddere, væsler, hermeliner og jærv. Som mange andre mustelider er de kødædende pattedyr, der lever af en række forskellige byttedyr som gnavere, småfugle og insekter. En af de mere udbredte arter, som de lever af, er egern: både røde egern og grå egern er potentiel føde for den søde, men vilde skovmår.
Men inden for skovmårens udbredelse (over store dele af Europa) er røde egern den indfødte art, og grå egern er invasive, idet de stammer fra Nordamerika. På grund af det langvarige forhold mellem røde egern og skovmårer har de udviklet sig sammen: mest markant ved at røde egern har ændret sig til en livsstil, der hovedsageligt foregår i træer, og ved at undgå jorden så meget som muligt. Grå egern har imidlertid ikke haft den evolutionære historie til at lære denne lektie og er nem mad for en smart fyrremarder. I de områder, hvor der er blevet bevaret eller genindført skovmårer, bekæmper de således aktivt den invasive bestand af grå egern, hvilket igen styrker den indfødte bestand af røde egern ved at mindske konkurrencen. Den koevolutionære forbindelse mellem røde egern og skovmårer er afgørende for bekæmpelsen af den invasive art.

Vært-parasit-koevolution
På samme måde som rovdyr og bytte-koevolutionen gennemgår patogene arter og deres uheldige værter også en slags ‘våbenkapløb’. Parasitterne må hele tiden udvikle nye måder at inficere og overføre til værterne på, samtidig med at værterne udvikler nye metoder til at modstå og undgå de inficerende arter. Denne spiralformede kamp mellem evolutionære kræfter kaldes “Red Queen-hypotesen”, der blev formuleret i 1973 af Leigh Van Valen og bruges til at beskrive mange andre former for koevolution. Navnet stammer fra Lewis Carrolls “Gennem spejlet” og især et citat:
“Her skal man løbe alt hvad man kan for at blive på samme sted”.
Citatet henviser til, hvordan arter hele tiden må tilpasse sig og reagere på andre arters udvikling for at blive ved med at eksistere og forhindre udryddelse. Arter, der forbliver statiske og holder op med at udvikle sig, vil uundgåeligt uddø i takt med, at verden omkring dem ændrer sig.
Mimicry
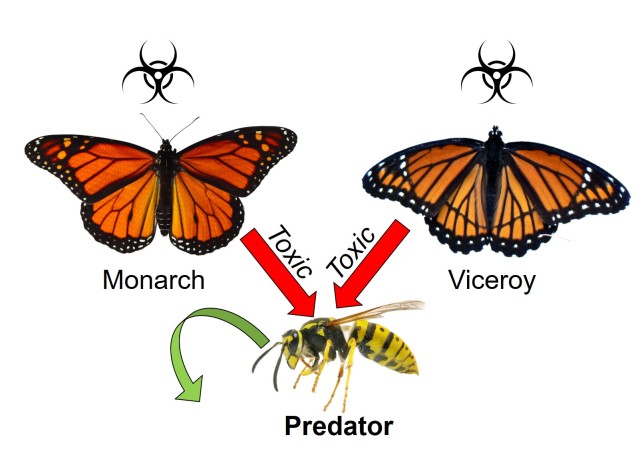
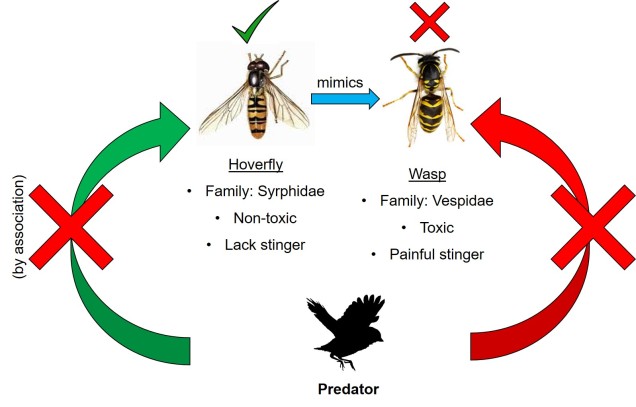
Der findes masser af andre mærkelige og unikke mekanismer for samudvikling i naturen. En af dem er mimikry, den proces, hvorved en art forsøger at ligne en anden for at beskytte sig selv. Den mest ikoniske gruppe, der er kendt for dette, er sommerfugle: mange arter deler lignende farvemønstre og kropsformer som efterlignere, selv om de evolutionært set kan være meget forskellige. Afhængigt af efterligningens art kan efterligning klassificeres i to brede kategorier. I begge tilfælde er den oprindelige “referenceart” giftig eller usmagelig for rovdyr og bruger en form for farvesignal for at kommunikere dette: tænk på biernes og hvepsenes klare gule farver eller mariehønenes røde farver. Der, hvor de to kategorier ændrer sig, er i arten af den “efterlignende” art.
Müllerisk mimik
Hvis den efterlignende art også er giftig eller uspiselig, kalder vi dette Müllerisk mimik (efter Johann Friedrich Theodor Müller). Ved at dele de samme farvemønstre og ved at begge er giftige øger de to efterlignende arter potentialet for, at rovdyr kan lære signalet. Hvis et rovdyr spiser en af arterne, vil det forbinde det pågældende farvemønster med giftighed, og ingen af arterne er så tilbøjelige til at blive angrebet i fremtiden. I den forstand er der tale om et kooperativt coevolutionært forhold mellem de to fysisk ensartede arter.
Batesiansk mimik
Derimod er det muligt, at efterligneren faktisk ikke er giftig eller usmagelig, men blot kopierer en giftig art. Dette kaldes Batesian mimicry (efter Henry Walter Bates) og indebærer, at den efterlignende art er afhængig af, at rovdyrene har lært at forbinde farve og toksicitet gennem “referencearten”. Selv om den efterlignende art ikke er giftig, er den i bund og grund en efterligner af det hårde evolutionære arbejde, der allerede er udført af den faktisk giftige art. I dette tilfælde er det koevolutionære forhold mere parasitært, da efterligneren drager fordel af “referencearten”, men den gengælder ikke denne fordel.
Arternes koevolution og betydningen af artsinteraktioner
Der er utallige andre artsinteraktioner, som kan drive koevolutionære relationer i naturen. Disse kan omfatte forskellige former for symbiose eller forskellige arters reaktion på økosystemingeniører: dvs. arter, der kan ændre og forme miljøet omkring dem (f.eks. koraller i revsystemer). For at forstå, hvordan en art udvikler sig i sit miljø, må man derfor tage hensyn til, hvordan mange andre lokale arter også udvikler sig og reagerer på deres egen måde.