
Isolering og karakterisering af pterosinderivater fra P. aquilinum
Pterosinderivater blev isoleret fra hele planter af P. aquilinum. Til undersøgelse af de fytokemiske bestanddele fra bregnerne blev et vandekstrakt successivt partitioneret med ethylacetat (EtOAc) og n-butanol (n-BuOH) (fig. 1). Gentagen kolonnekromatografi af den EtOAc-opløselige fraktion resulterede i isolering af ni derivater, herunder (2R)-pterosin B (1), pterosin Z (2), (2S)-pterosin P (3), (3R)-pterosin D (4), (2S)-pterosin A (5), (2S,3R)-pterosin C (6), (2R,3R)-pterosin C (7), (2R)-pterosid B (8) og pterosid Z (9), med en renhed på over 97 % (Fig. 2). Den gentagne kolonnekromatografiske separation af den n-BuOH-opløselige fraktion resulterede i isolering af seks derivater, herunder (2S)-pterosid A (10), (2S)-pterosid A2 (11), (2S,3R)-pterosid C (12), (2R,3R)-pterosid C (13), (3S)-pterosid D (14) og (2S)-pterosid P (15), med en renhed på over 97 %. Forbindelsernes strukturer blev identificeret ved analyse af spektraldata, herunder MS, 1D- og 2D-NMR (Supplerende information 1).

Strukturer af pterosinforbindelser 1-15xxx
Inhiberende aktivitet af pterosinderivater mod BACE1, AChE, og BChE
For at evaluere anti-AD-potentialet blev den inhiberende aktivitet af hver pterosinforbindelse mod BACE1 og cholinesteraser evalueret ved respektive in vitro-inhibitionsassays (tabel 1). Alle testede pterosinderivater viste koncentrationsafhængige hæmmende aktiviteter mod BACE1 med et interval af IC50-værdier (halvmaksimale hæmmende koncentration) på 9,74-94,4 μM, med undtagelse af (2S)-pterosin A og (2S)-pterosid P, der var inaktive ved de testede koncentrationer, sammenlignet med IC50 af quercetin, der blev anvendt som positiv kontrol, som var 18,8 μM. Den hæmmende virkning af de stærkeste inhibitorer var i rækkefølge af (2R,3R)-pterosid C, (3S)-pterosid D, (2R)-pterosid B, (2S,3R)-pterosin C, (2R,3R)-pterosin C, (2S,3R)-pterosid C og (2R)-pterosin B med IC50-værdier på 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 og 29,6 μM, henholdsvis. Vi testede efterfølgende de hæmmende potentialer af pterosinderivaterne mod AChE. Alle de testede forbindelser viste betydelige AChE-hæmmende aktiviteter med IC50-værdier i intervallet 2,55-110 μM sammenlignet med IC50-værdien mod AChE for berberin, der blev anvendt som positiv kontrol, som var 0,39 μM. De pterosinforbindelser, der udviste den stærkeste hæmmende aktivitet over for AChE, var (2R)-pterosid B, (2R,3R)-pterosid C, (2S,3R)-pterosid C, (2S,3R)-pterosin C og (2R)-pterosin B med IC50-værdier på henholdsvis 2,55, 3,77, 9,17, 12,8 og 16,2 μM. Endelig testede vi den hæmmende kapacitet af pterosinderivaterne mod BChE. Alle de testede pterosinforbindelser viste hæmmende aktivitet mod BChE med IC50-værdier, der varierede fra 5,29 til 119 μM, med undtagelse af (3R)-pterosin D, der var inaktivt ved de testede koncentrationer, sammenlignet med berberins IC50 mod BChE, som var 3,32 μM. De pterosinforbindelser, der udviste den stærkeste hæmmende aktivitet over for BChE, var (2R,3R)-pterosid C og pterosid Z med IC50-værdier på henholdsvis 5,29 og 5,31 μM.
Samlet set udviste de fleste af de testede pterosinderivater signifikante hæmmende aktiviteter mod BACE1, AChE og BChE samtidig. Tilstedeværelsen af den ekstra 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolgruppe som i pterosidderivater øgede de hæmmende aktiviteter mod enzymerne betydeligt. Desuden mindskede tilstedeværelsen af den ekstra hydroxymethylgruppe på position 2 i indanonringen i (2R)-pterosin B som i (2S)-pterosin A eller methylgruppen som i (3R)-pterosin D og pterosin Z de hæmmende aktiviteter over for enzymerne. Især tilstedeværelsen af hydroxymethylgruppen på position-5 i indanonringen som i (2S)-pterosin P mindskede den hæmmende aktivitet mod BACE1.
Kinetiske parametre for enzymhæmning af pterosinderivater
I et forsøg på at forklare den enzymatiske hæmningsmåde af pterosinderivater udførte vi en kinetisk analyse af BACE1 og kolinesteraser for repræsentative inhibitorer (Tabel 2, Supplerende information 2). En lav Ki (hæmningskonstant) indikerer en tættere enzymbinding og en mere effektiv inhibitor. Samlet set korrelerede forbindelsernes Ki-værdier med de respektive IC50-værdier. BACE1-hæmning af forbindelserne (2R,3R)-pterosid C, (3S)-pterosid D og (2R,3R)-pterosin C var af blandet type med Ki værdier på 12,6, 16,5 og 27.6 µM, mens hæmning af (2R)-pterosid B, (2S,3R)-pterosin C og (2R)-pterosin B var ikke-kompetitiv med Ki-værdier på henholdsvis 23,1, 33,8 og 38,3 µM. AChE-hæmning af (2R)-pterosid B, (2R,3R)-pterosid C, (2R)-pterosin B, (2S,3R)-pterosin C og (3S)-pterosid D var af blandet type med Ki-værdier på 4.89, 8,13, 12,1, 16,3 og 23,1 µM, mens (2R,3R)-pterosin C var en ikke-konkurrerende typehæmmer med en Kiværdi på 29,6 µM. BChE-hæmning af (2R,3R)-pterosin C, (2R,3R)-pterosid C, (3S)-pterosid D og (2R)-pterosid B var af blandet type med Ki-værdier på 4,77, 9,62, 19.7 og 22,6 µM, mens (2S,3R)-pterosin C og (2R)-pterosin B var ikke-kompetitive inhibitorer med Ki værdier på henholdsvis 29,9 og 53,5 µM. Disse resultater tyder således på, at specifikke pterosinderivater kan være effektive BACE1-, AChE- og BChE-inhibitorer.
Molekylære docking-simuleringer for BACE1, AChE og BChE
Der findes adskillige krystalstrukturer for BACE1 og kolinesteraser. Vi udvalgte menneskelige PDB’er baseret på wild-type-strukturer, samkrystalliserede ligander og opløsninger af strukturerne. Røntgenkrystalstrukturer af BACE1 kompleks med QUD (PDB-kode: 2WJO, opløsning: 2,5 Å)33, AChE kompleks med E2020 (PDB-kode: 4EY7, opløsning: 2,35 Å)34 og BChE kompleks med 3F9 (PDB-kode: 4TPK, opløsning: 2,70 Å)35 blev udvalgt med henblik på docking. Oprindeligt blev QUD, E2020 og 3F9 ekstraheret fra krystalstrukturer og redocket ind i de aktive steder for henholdsvis BACE1, AChE og BChE. Derefter blev (2R,3R)-pterosid C, (3S)-pterosid D, (2R,3R)-pterosin C, (2R)-pterosid B, (2S,3R)-pterosin C og (2R)-pterosin B med den kendte hæmningsmekanisme mod BACE1, AChE og BChE docket med henblik på at bestemme deres SAR. Dockingresultaterne er opsummeret i tabel 3. SAR for de udvalgte blandede og ikke-kompetitive BACE1-, AChE- og BChE-hæmmere af blandet type gjorde det muligt at evaluere pterosinernes nye bindingsstedsbestemte hæmning af enzymerne.
Vores dockingmodus for E2020 var i overensstemmelse med den eksperimentelt bestemte bindingsmodus, der tidligere er rapporteret med rekombinant human AChE (rhAChE) (Supplerende oplysninger 3)34. Root-mean-square-afvigelsen (RMSD) mellem krystal- og dockingkonformationerne af E202020 var 0,54 Å, hvilket tyder på pålideligheden af vores dockingopsætning med hensyn til at reproducere den eksperimentelle bindingsmodus. Desuden førte E2020’s dockingtilstand til en lignende interaktion som rhAChE-E202020. I vores undersøgelse blev vandmolekyler fjernet fra krystalstrukturen under docking; derfor blev vandmedierede interaktioner ikke analyseret i den foreliggende undersøgelse. På samme måde var de dockingmodes af QUD og 3F9 i overensstemmelse med de tilgængelige eksperimentelle data for henholdsvis BACE1 33 og BChE35 (Supplerende oplysninger 3). RMSD’erne mellem krystalkonformationerne og de dokkede konformationer af QUD og 3F9 var henholdsvis 0,46 og 0,60 Å. Endvidere var bindingsstederne for pterosinhæmmerne i overensstemmelse med en tidligere dockingundersøgelse, der involverede BACE1, AChE og BChE38. Undersøgelsen anvendte imidlertid Tetronarce californica AChE (PDB-kode: 1ACJ), som indeholder lidt forskellige restnumre end human AChE på grund af variationer i deres sekvenser.
BACE1-dokning
Baseret på hæmningstype og aktivitet blev (2R,3R)-pterosid C og (2R)-pterosid B udvalgt til at demonstrere de dokkede tilstande for henholdsvis blandede typer og ikke-kompetitive BACE1-hæmmere. Figur 3a og b viser dockingmodellerne for henholdsvis (2R,3R)-pterosid C og (2R)-pterosid B. Interaktionerne mellem de dockingmodellerede forbindelser i det aktive sted for BACE1 er vist i figur 4.

Molekylære dockingmodeller for a BACE1-inhibitoren af blandet type (2R,3R)-pterosid C (blå farve) og b den ikke-kompetitive BACE1-inhibitor (2R)-pterosid B (brun farve). De dockingpositioner er overlejret på røntgenkrystalstrukturen af QUD (gul farve) (PDB-kode: 2WJO). BACE1, rester på det aktive sted og forbindelser er vist ved henholdsvis bånd-, linje- og stavmodeller. Farverne på de stiplede linjer forklarer typerne af de forskellige interaktioner: hydrogenbindingsinteraktioner (grøn), hydrofobiske interaktioner (pink) og π-sigma-interaktioner (lilla). BACE1 β-site amyloidprækursorprotein-spaltningsenzym 1

Ligandinteraktionsdiagram for a QUD, b (2R,3R)-pterosid C, c (3S)-pterosid D, d (2R,3R)-pterosin C, e (2R)-pterosid B, f (2S,3R)-pterosin C og g (2R)-pterosin B i det aktive sted for BACE1. Farverne på de stiplede linjer forklarer typerne af de forskellige interaktioner: hydrogenbindingsinteraktioner (grøn), hydrofobiske interaktioner (pink) og π-sigma-interaktioner (lilla). BACE1 β-site amyloid precursor protein cleaving enzyme 1
Den dockede pose af QUD udviste en bindingsenergi (B.E.) på -7,59 kcal/mol. Som vist i fig. 4a viste NH2-gruppen på ligandens quinazolinring to hydrogenbindinger med CO-grupperne i Asp32 og Asp228 i afstande på henholdsvis 1,86 og 2,17 Å. En tredje hydrogenbinding blev observeret mellem ligandens anden NH-gruppe og CO-gruppen af Gly230 i en afstand på 2,16 Å. Leu30, Tyr71, Phe108 og Val332 formidlede de hydrofobiske interaktioner. Figur 4b-d viser de dockingpositioner af henholdsvis (2R,3R)-pterosid C, (3S)-pterosid D og (2R,3R)-pterosin C (BACE1-hæmmere af blandet type). De blev placeret i den bindingslomme, der er beklædt af Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 og Thr231. I henhold til deres aktivitetsniveauer udviste (2R,3R)-pterosid C (IC50 = 9,74 µM), (3S)-pterosid D (IC50 = 10,7 µM) og (2R,3R)-pterosin C (IC50 = 26,2 µM) en B.E. på henholdsvis -7,27, -6,93 og -4,84 kcal/mol. (2R,3R)-Pterosid C udviste en højere potens end (2R,3R)-Pterosin C på grund af eksistensen af en ekstra 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolgruppe, som dannede tre hydrogenbindinger (Fig. 4b). OH-gruppen af hydroxymethyl og 3-OH-gruppen af tetrahydro-pyran-triolringen viste to hydrogenbindinger med CO-gruppen af Asp228 i afstande på henholdsvis 2,63 og 2,12 Å. Desuden viste 3-OH-gruppen en hydrogenbinding med Thr231 i en afstand på 1,89 Å. I tilfælde af (3S)-pterosid D (Fig. 4c) ændrede tilstedeværelsen af 2,2-dimethylgruppen ved indanonringen lidt på bindingsinteraktionerne sammenlignet med (2R,3R)-pterosid C. 3-OH-gruppen af tetrahydro-pyran-triolringen viste kun en enkelt hydrogenbinding med Asp228 i en afstand på 1,93 Å. Indanonringens 3-OH-gruppe udviste imidlertid en yderligere hydrogenbinding med Ile126 i en afstand på 2,34 Å. Disse interaktioner sænkede lidt aktiviteten af (3S)-pterosid D sammenlignet med (2R,3R)-pterosid C. Dockinginteraktionerne for (2R,3R)-pterosin C, der vises i fig. 4d, viser, at 3-OH-gruppen i indanonringen er bundet til Ser36 og Asn37 via to hydrogenbindinger i afstande på henholdsvis 1,83 og 2,07 Å. Ala39, Val69, Trp76, Ile118 og Arg128 var involveret i hydrofobiske interaktioner, mens Val69 udviste en π-sigma-interaktion.
De dockede positurer af (2R)-pterosid B, (2S,3R)-pterosin C og (2R)-pterosin B (ikke-kompetitive BACE1-hæmmere) er vist i henholdsvis Fig. 4e-g. De blev docket ind i det hulrum, der er omsluttet af Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 og Ile126. I overensstemmelse med deres aktivitetsværdier udviste (2R)-pterosid B (IC50 = 18,0 µM), (2S,3R)-pterosin C (IC50 = 23,1 µM) og (2R)-pterosin B (IC50 = 29,6 µM) en B.E. på henholdsvis -6,16, -5,07 og -4,64 kcal/mol. Som vist i fig. 4e udviste (2R)-pterosid B højere aktivitet end (2S,3R)-pterosin C og (2R)-pterosin B på grund af tilstedeværelsen af en yderligere 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol-gruppe, som viste fire hydrogenbindingsinteraktioner. Der blev observeret to hydrogenbindinger mellem 4-OH-gruppen i tetrahydro-pyran-triolringen og NH- og CO-grupperne i Asn37 med en afstand på henholdsvis 2,48 og 2,19 Å. Endvidere viste 3- og 5-OH-grupperne yderligere to hydrogenbindinger med Ile126 og Trp76 i afstande på henholdsvis 2,19 og 2,46 Å. (2S,3R)-Pterosin C viste en lidt bedre aktivitet end (2R)-Pterosin B på grund af tilstedeværelsen af en ekstra OH-gruppe på position-3 af indanonringen, som dannede en hydrogenbinding med Lys107 i en afstand på 2,12 Å (Fig. 4f). De andre interaktioner lignede dem for (2R)-pterosin B. Som vist i fig. 4g viste (2R)-pterosin B hydrofobiske interaktioner med Val69, Tyr76 og Phe108.
AChE-dokning
(2R)-Pterosid B og (2R,3R)-pterosin C blev udvalgt som repræsentanter for at demonstrere de dockingmodes af henholdsvis blandede type og ikke-kompetitive AChE-hæmmere på grund af deres aktiviteter og type AChE-hæmning. Figur 5a og b illustrerer dockingmodellerne for henholdsvis (2R)-pterosid B og (2R,3R)-pterosin C. Interaktionerne mellem de dockingmodellerede forbindelser inden for AChE’s aktive sted er vist i figur 6.

Molekylære dockingmodeller for a den blandede type AChE-inhibitor (2R)-pterosid B (blå farve) og b den ikke-kompetitive AChE-inhibitor (2R,3R)-pterosin C (brun farve). De dockingpositioner er overlejret på røntgenkrystalstrukturen af E2020 (gul farve) (PDB-kode: 4EY7). AChE, rester på det aktive sted og forbindelser er vist med henholdsvis bånd-, linje- og stavmodeller. Farverne på de stiplede linjer forklarer typerne af de forskellige interaktioner: hydrogenbindingsinteraktioner (grøn) og hydrofobiske interaktioner (pink). AChE acetylcholinesterase

Ligandinteraktionsdiagram for a E2020, b (2R)-pterosid B, c (2R,3R)-pterosid C, d (2S,3R)-pterosin C, e (2R)-pterosin B, f (3S)-pterosid D og g (2R,3R)-pterosin C i det aktive sted for AChE. Farverne på de stiplede linjer forklarer typerne af de forskellige interaktioner: hydrogenbindingsinteraktioner (grøn), hydrofobiske interaktioner (pink) og π-sigma-interaktioner (lilla). AChE acetylcholinesterase
Den dockede positur af E2020 viste en B.E. på -10,28 kcal/mol. Som illustreret i Fig. 6a dannede CO-gruppen i indanonringen en hydrogenbinding med NH-gruppen af Phe295 i en afstand på 1,70 Å. Trp286 og Tyr341 var involveret i π-sigma-interaktioner, mens Trp86, Trp286, Tyr337, Phe338 og Tyr341 medierede hydrofobiske interaktioner. Figur 6b-f viser de dockede positurer af henholdsvis (2R)-pterosid B, (2R,3R)-pterosid C, (2S,3R)-pterosin C, (2R)-pterosin B og (3S)-pterosid D (AChE-hæmmere af blandet type). De blev anbragt i det aktive sted omgivet af Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 og Tyr341. I overensstemmelse med deres aktivitetsværdier er (2R)-pterosid B (IC50 = 2,55 µM), (2R,3R)-pterosid C (IC50 = 3,77 µM), (2S,3R)-pterosin C (IC50 = 12.8 µM), (2R)-pterosin B (IC50 = 16,2 µM) og (3S)-pterosid D (IC50 = 27,4 µM) udviste en B.E. på henholdsvis -7,90, -7,49, -6,03, -5,76 og -4,91 kcal/mol. (2R)-Pterosid B udviste en højere potens end (2S,3R)-Pterosin C og (2R)-Pterosin B på grund af tilstedeværelsen af en ekstra 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol-gruppe, som etablerede tre hydrogenbindingsinteraktioner (Fig. 6b). OH-gruppen af hydroxymethyl og 3-OH-gruppen af tetrahydro-pyran-triolringen udviste hydrogenbindinger med CO-gruppen af Asn87 i afstande på henholdsvis 2,23 og 2,28 Å. Endvidere dannede 4-OH-gruppen en hydrogenbinding med CO-gruppen af Trp86 i en afstand på 2,13 Å. I tilfælde af (2R,3R)-pterosid C (fig. 6c) viste methylgruppen i position-2 af indanonringen ikke en hydrofob interaktion med Tyr341 og udviste således forholdsvis lavere aktivitet end (2R)-pterosid B. Tyr341 opretholdt imidlertid den hydrofobiske interaktion med den anden del af indanonringen som vist i (2R)-pterosid B. Den højere aktivitet af (2S,3R)-pterosin C end (2R)-pterosin B blev tilskrevet eksistensen af en ekstra OH-gruppe i position-3 af indanonringen, som dannede en hydrogenbinding med Phe295 i en afstand på 1,82 Å (fig. 6d). De resterende interaktioner var sammenlignelige med (2R)-pterosin B-interaktioner. Som vist i Fig. 6e dannede OH-gruppen af hydroxyethylgruppen på position-6 af indanonringen en hydrogenbinding med Tyr124 i en afstand på 2,48 Å. Tyr124, Trp286, Tyr337, Phe338 og Tyr341 bidrog til de hydrofobiske interaktioner. Sammenlignet med (2R)-pterosid B og (2R,3R)-pterosid C udviste (3S)-pterosid D uensartede bindingsinteraktioner på grund af tilstedeværelsen af 2,2-dimethylgruppen ved indanonringen (fig. 6f). 2,2-dimethylgruppen bidrog væsentligt til den særskilte dockingposition af (3S)-pterosid D. 5-OH-gruppen i tetrahydro-pyran-triolringen dannede en hydrogenbinding med Asp74 i en afstand på 1,96 Å. CO-gruppen i indanonringen udviste en hydrogenbinding med Tyr72 i en afstand af 2,97 Å. Disse interaktioner forklarede den lave aktivitet af (3S)-pterosid D.
Den dockingposition af (2R,3R)-pterosin C (ikke-kompetitiv AChE-hæmmer) er vist i fig. 6g. (2R,3R)-Pterosin C (IC50 = 23,2 µM) viste en B.E. på -5,01 kcal/mol. Bindingslommen for (2R,3R)-pterosin C omfattede Tyr72, Trp286, Ser293, Phe297 og Tyr341, med to hydrogenbindingsinteraktioner. En af hydrogenbindingerne blev dannet mellem 3-OH-gruppen af indanonringen og CO-gruppen af Ser293 i en afstand på 2,03 Å. Den anden hydrogenbinding blev observeret mellem OH-gruppen af hydroxyethylgruppen, der er til stede i position-6 af indanonringen, og Tyr72 i en afstand på 1,94 Å. Residuer som Tyr72, Trp286, Phe297 og Tyr341 deltog i hydrofobiske interaktioner.
BChE-dokning
I betragtning af aktivitetsniveauerne og typen af BChE-hæmning blev (2R,3R)-pterosid C og (2S,3R)-pterosin C udvalgt til at demonstrere de dokkede tilstande for henholdsvis blandede typer og ikke-kompetitive BChE-hæmmere. Figur 7a og b illustrerer dockingmodellerne for henholdsvis (2R,3R)-pterosid C og (2S,3R)-pterosin C. Interaktionerne mellem de dockede forbindelser i det aktive BChE-sted er vist i figur 8.
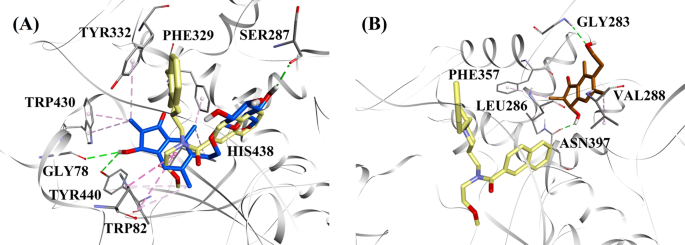
Molekylære dockingmodeller for a BChE-inhibitoren af blandet type (2R,3R)-pterosid C (blå farve) og b den ikke-kompetitive BChE-inhibitor (2S,3R)-pterosin C (brun farve). De dockingpositioner er overlejret på røntgenkrystalstrukturen af 3F9 (gul farve) (PDB-kode: 4TPK). BChE, rester på det aktive sted og forbindelser er vist med henholdsvis bånd-, linje- og stavmodeller. Farverne på de stiplede linjer forklarer typerne af de forskellige interaktioner: hydrogenbindingsinteraktioner (grøn) og hydrofobiske interaktioner (pink). BChE butyrylcholinesterase
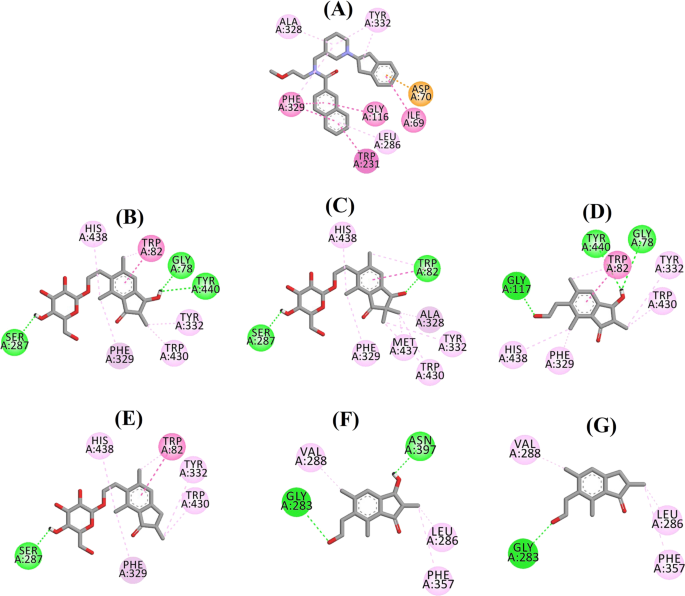
Ligandinteraktionsdiagram for a 3F9, b (2R,3R)-pterosid C, c (3S)-pterosid D, d (2R,3R)-pterosin C, e (2R)-pterosid B, f (2S,3R)-pterosin C og g (2R)-pterosin B i det aktive sted for BChE. Farverne på de stiplede linjer forklarer typerne af de forskellige interaktioner: hydrogenbindingsinteraktioner (grøn), hydrofobiske interaktioner (pink) og π-anioninteraktioner (gylden). BChE butyrylcholinesterase
Den dockede positur af 3F9 viste en B.E. på -8,49 kcal/mol. Som vist i fig. 8a var hydrofobiske interaktioner hovedsageligt ansvarlige for ligandbindingen. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 og Tyr332 stod for de hydrofobiske interaktioner, mens Asp70 udviste en π-anioninteraktion. Figur 8b-e illustrerer de dockingpositioner for henholdsvis (2R,3R)-pterosid C, (3S)-pterosid D, (2R,3R)-pterosin C og (2R)-pterosid B (BChE-hæmmere af blandet type). Deres bindingslomme var sammensat af Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 og Tyr440. I overensstemmelse med deres aktivitetsniveauer udviste (2R,3R)-pterosid C (IC50 = 5,29 µM), (3S)-pterosid D (IC50 = 19,3 µM), (2R,3R)-pterosin C (IC50 = 20,3 µM) og (2R)-pterosid B (IC50 = 62,0 µM) en B.E. på henholdsvis -7,23, -6,59, -6,52 og -4,38 kcal/mol. Som vist i fig. 8b viste 3-OH-gruppen i indanonringen af (2R,3R)-pterosid C to hydrogenbindinger med Gly78 og Tyr440 i afstande på henholdsvis 2,87 og 2,89 Å. I tilfældet med (3S)-pterosid D påvirkede 2,2-dimethylgruppen på indanonringen bindingsinteraktionerne for 3-OH-gruppen (fig. 8c). 3-OH-gruppen dannede kun én hydrogenbinding med Trp82 i en afstand på 2,94 Å, hvilket resulterede i en forholdsvis lavere aktivitet af (3S)-pterosid D end (2R,3R)-pterosid C. Som vist i Fig. 8d formåede (2R,3R)-pterosin C ikke at danne en hydrogenbinding med Ser287 på grund af fraværet af en 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol-gruppe. Det gav derfor en lavere aktivitet end (2R,3R)-pterosid C. Manglen på OH-gruppen i position-3 af indanonringen viste sig at være ansvarlig for den meget lave aktivitet af (2R)-pterosid B (Fig. 8e), som ikke formåede at danne hydrogenbindinger med Gly78 og Tyr440 i lighed med (2R,3R)-pterosid C.
De dockede positurer af (2S,3R)-pterosin C og (2R)-pterosin B (ikke-kompetitive BChE-hæmmere) er vist i henholdsvis Fig. 8f, g. Disse dockingpositioner var indeholdt i det hulrum, der er omsluttet af Gly283, Leu286, Val288, Phe357 og Asn397. I henhold til deres aktivitetsniveauer udviste (2S,3R)-pterosin C (IC50 = 44,3 µM) og (2R)-pterosin B (IC50 = 48,1 µM) en B.E. på henholdsvis -5,40 og -5,06 kcal/mol. (2S,3 R)-Pterosin C var mere potent end (2R)-pterosin B på grund af tilstedeværelsen af en ekstra OH-gruppe på position-3 af indanonringen, som dannede en hydrogenbinding med CO-gruppen af Asn397 i en afstand på 2,05 Å (Fig. 8f). Andre interaktioner viste sig at ligne (2R)-pterosin B. Som vist i fig. 8g dannede OH-gruppen i hydroxyethylgruppen i indanonringens position-6 en hydrogenbinding med Gly283 i en afstand på 2,40 Å. Resterne Leu286, Val288 og Phe357 deltog i hydrofobiske interaktioner.
Mixed-type inhibitorer binder både til det frie enzym og enzym-substratkomplekset, hvilket indikerer, at disse forbindelser kan binde til det katalytiske sted for hvert tilsvarende enzym. Ikke-kompetitive inhibitorer binder til det allosteriske sted i det frie enzym eller enzym-substratkomplekset. En nylig undersøgelse tyder på, at kompetitive, blandede og ikke-kompetitive inhibitorer optager forskellige steder i bindingslommerne for BACE1, AChE og BChE38. Under docking til evaluering af pterosinderivaternes hæmmende mekanisme blev forbindelsernes bindingssteder defineret i henhold til deres type hæmning. Dockingresultaterne viste, at bindingsstederne for blandede typer og ikke-kompetitive inhibitorer for BACE1, AChE og BChE delvis overlapper hinanden på hvert tilsvarende aktive sted og var i overensstemmelse med en tidligere rapport38.
BBB-permeabilitet
PAMPA-BBB, et in vitro kunstigt membranpermeabilitetsassay for BBB, er et af de mest pålidelige fysisk-kemiske screeningsværktøjer i den tidlige fase af opdagelsen af lægemidler rettet mod CNS40. PAMPA-BBB-systemet modellerer den transcellulære passive diffusion af kemikalier gennem BBB og måler udelukkende passive transportmekanismer via en kunstig lipidmembran på den effektive permeabilitet (Pe, cm/s). På grundlag af det mønster, der er fastlagt for forudsigelse af BBB-permeation, blev stofferne klassificeret i) “CNS+” (høj forudsagt BBB-permeation); Pe (10-6 cm/s) > 4,00, (ii) “CNS-” (lav forudsagt BBB-permeation); Pe (10-6 cm/s) < 2,00 og (iii) “CNS+/-” (usikker BBB-permeation); Pe (10-6 cm/s) fra 4,00 til 2,00. Følgelig udviste (2R)-pterosin B, (2S)-pterosin P og (2S)-pterosin A høj BBB-permeation med Pe-værdier på henholdsvis 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s og 6,26 × 10-6 cm/s (Tabel 4, Supplerende oplysninger 4). Pe-værdien af (2R)-pterosin B var 1,7 gange højere end Pe-værdien af CNS-medikamentet verapamil (Pe = 34,6 × 10-6 cm/s), som blev anvendt som positiv kontrol. (2S,3R)-Pterosin C og (2R,3R)-pterosin C viste en usikker BBB-permeation med Pe-værdier på henholdsvis 2,34 og 1,98. (2R,3R)-Pterosid C, (3S)-Pterosid D og (2R)-Pterosid B, som viste de mest potente BACE1- og cholinesterasehæmmende aktiviteter blandt de testede pterosinderivater, udviste en meget lav BBB-permeabilitet. Tilstedeværelsen af 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol-gruppen som i pterosidderivater resulterede i et bemærkelsesværdigt fald i BBB-permeabiliteten. Sammenlignet med (2R)-pterosin B reducerede den yderligere tilstedeværelse af OH-gruppen i position-3 af indanonringen som i pterosin C, hydroxymethylgruppen i position-2 af indanonringen som i (2S)-pterosin A eller hydroxymethylgruppen i position-5 af indanonringen som i (2S)-pterosin P også BBB-permeabiliteten betydeligt. I betragtning af en usædvanlig høj BBB-permeabilitet og den betydelige hæmning af BACE1, AChE og BChE kan (2R)-pterosin B have potentiale til at udvise en stærk anti-AD-aktivitet.
Virkninger af (2R)-pterosin B og (2R,3R)-pterosid C på sekretionen af Aβ-peptider fra neuronale celler
For at undersøge funktionen af (2R)-pterosin B og (2R,3R)-pterosid C med hensyn til at nedsætte udskillelsen af Aβ fra neuronale celler, anvendte vi en murin neuroblastomcellelinje, der stabilt overudtrykker human APPswe. Cellelinjen er en cellulær model af AD, der er karakteriseret ved overdreven udskillelse af Aβ40 og Aβ42. Giftige amyloidoligomerer dannes af de to isoformer af Aβ-peptidet med forskellige længder. Aβ40 er den hyppigst forekommende Aβ-isoform i hjernen41 , mens Aβ42 stiger betydeligt med visse former for AD42. Sandwich ELISA af Aβ40 viste, at (2R)-pterosin B signifikant reducerede mængden af Aβ40-peptid, der blev udskilt fra neuroblastomcellerne til medierne, med op til 50 % ved 500 μM (P < 0,01) (Fig. 9a) (Fig. 9a). Tilsvarende faldt udskillelsen af Aβ42-peptidet fra neuroblastomcellerne signifikant i tilstedeværelsen af 500 µM (2R,3R)-pterosid C (P < 0,05) (Fig. 9b) (Fig. 9b). Det kan konkluderes, at (2R)-pterosin B og (2R,3R)-pterosid C signifikant nedsatte sekretionen af Aβ-peptider fra neuroblastomceller ved en koncentration på 500 μM.

a Virkning af (2R)-pterosin B på sekretionen af Aβ-peptider. APPswe-secernende neuroblastomceller blev behandlet med 12, 60, 250 eller 500 µM (2R)-pterosin B i 24 timer, og konditionerede medier blev opsamlet i tilstedeværelse af proteasehæmmer. β-SI (10 µM) blev anvendt som positiv kontrol. Negativ kontrol omfattede celler, der var dyrket i fravær af testforbindelser. Kvantitativ analyse af udskilt Aβ40 og Aβ42 i de konditionerede medier blev udført ved hjælp af sandwich-ELISA. Middelværdierne ± SD fra tre uafhængige eksperimenter er vist. De sekreterede Aβ40-peptider faldt signifikant i tilstedeværelsen af 500 µM (2R)-pterosin B. CON negativ kontrol, β-SI β-sekretaseinhibitor III, Aβ β-amyloid. *P < 0,05, **P < 0,01. b Effekt af (2R,3R)-pterosid C på sekretionen af Aβ-peptider. APPswe-secernende neuroblastomceller blev behandlet med 12, 60, 125, 250 eller 500 µM (2R,3R)-pterosid C i 24 timer, og konditionerede medier blev opsamlet i tilstedeværelse af proteasehæmmer. β-SI (10 µM) blev anvendt som positiv kontrol. Negativ kontrol omfattede celler, der var dyrket i fravær af testforbindelser. Kvantitativ analyse af udskilt Aβ40 og Aβ42 i de konditionerede medier blev udført ved hjælp af sandwich-ELISA. Middelværdierne ± SD fra tre uafhængige eksperimenter er vist. Det sekreterede Aβ42-peptid faldt signifikant i tilstedeværelsen af 500 µM (2R,3R)-pterosid C. CON negativ kontrol, β-SI β-sekretaseinhibitor III, Aβ β-amyloid. *P < 0.05
Cytotoksicitet af pterosinderivater baseret på MTT-assay43
Samlet set udviste pterosinderivater en ubetydelig cytotoksicitet over for forskellige normale og kræftcellelinjer, såsom SH-SY5Y humant neuroblastom, C6 gliaceller fra rotter, NIH3T3 fibroblaster fra musebryoner og B16F10 musemelanom med LD50-værdier på over 0.5 mM (Supplerende oplysninger 5). Især (2R)-pterosin B, (3R)-pterosin D, (2S)-pterosin P, (2S)-pterosid A, (2R)-pterosid B og (2R,3R)-pterosid C viste ingen cytotoksicitet mod de testede cellelinjer med LD50-værdier på over 5 mM. Det er interessant, at flere pterosiner viste relative antiproliferative virkninger mod SH-SY5Y-neuronale celler sammenlignet med C6-gliaceller af mesenchymal oprindelse. De nuværende resultater var i overensstemmelse med en tidligere rapport, der tyder på, at pterosinderivater ikke er toksiske for mennesker26.