

Bidraget fra en saltbro til den samlede stabilitet til et proteins foldede tilstand kan vurderes ved hjælp af termodynamiske data indsamlet fra mutagenesestudier og kernemagnetiske resonansteknikker. Ved hjælp af et muteret pseudo-wild-typeprotein, der specifikt er muteret til at forhindre udfældning ved høj pH, kan saltbroens bidrag til den samlede frie energi for den foldede proteintilstand bestemmes ved at udføre en punktmutation, der ændrer og dermed bryder saltbroen. Det blev f.eks. konstateret, at der findes en saltbro i T4-lysozym mellem asparaginsyre (Asp) ved rest 70 og en histidin (His) ved rest 31 (figur 3). Der blev foretaget stedbestemt mutagenese med asparagin (Asn) (figur 4), hvorved der blev opnået tre nye mutanter: Asp70Asn His31 (Mutant 1), Asp70 His31Asn (Mutant 2) og Asp70Asn His31Asn (dobbeltmutant).

Når mutanterne er blevet etableret, kan der anvendes to metoder til at beregne den frie energi, der er forbundet med en saltbro. Den ene metode indebærer observation af smeltetemperaturen for wild-type proteinet i forhold til smeltetemperaturen for de tre mutanter. Denatureringen kan overvåges ved hjælp af en ændring i cirkulær dichroisme. En reduktion i smeltetemperaturen indikerer en reduktion i stabiliteten. Dette kvantificeres ved hjælp af en metode, der er beskrevet af Becktel og Schellman, hvor forskellen i fri energi mellem de to stoffer beregnes ved hjælp af ΔTΔS. Der er visse problemer med denne beregning, og den kan kun anvendes med meget nøjagtige data. I eksemplet med T4-lysozym var ΔS af pseudo-wild-typen tidligere blevet rapporteret ved pH 5,5, så den midterste temperaturforskel på 11 °C ved denne pH multipliceret med den rapporterede ΔS på 360 cal/(mol-K) (1,5 kJ/(mol-K)) giver en ændring i den frie energi på ca. -4 kcal/mol (-17 kJ/mol). Denne værdi svarer til den mængde fri energi, som saltbroen bidrager til proteinets stabilitet.
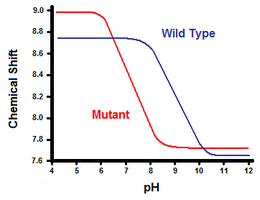
Den anden metode udnytter kernemagnetisk resonansspektroskopi til at beregne saltbroens frie energi. Der udføres en titrering, mens man registrerer det kemiske skift, der svarer til protonerne på det kulstof, der støder op til carboxylat- eller ammoniumgruppen. Midtpunktet af titreringskurven svarer til pKa, dvs. den pH-værdi, hvor forholdet mellem protonerede og deprotonerede molekyler er 1:1. I fortsættelse af eksemplet med T4-lysozym fås en titreringskurve ved observation af en forskydning af C2-protonen i histidin 31 (figur 5). Figur 5 viser forskydningen i titreringskurven mellem den vilde type og mutanten, hvor Asp70 er Asn. Den saltbro, der dannes, er mellem den deprotonerede Asp70 og den protonerede His31. Denne vekselvirkning forårsager den forskydning, der ses i His31’s pKa. I det udfoldede wild-type-protein, hvor saltbroen er fraværende, har His31 angiveligt en pKa på 6,8 i H20-buffere med moderat ionisk styrke. Figur 5 viser en pKa for vildtypen på 9,05. Denne forskel i pKa understøttes af His31’s interaktion med Asp70. For at opretholde saltbroen vil His31 forsøge at beholde sin proton så længe som muligt. Når saltbroen er afbrudt, som i mutanten D70N, skifter pKa tilbage til en værdi på 6,9, meget tættere på His31’s værdi i udfoldet tilstand.
Den forskel i pKa kan kvantificeres for at afspejle saltbroens bidrag til den frie energi. Ved hjælp af Gibbs frie energi: ΔG = -RT ln(Keq), hvor R er den universelle gaskonstant, T er temperaturen i kelvin, og Keq er ligevægtskonstanten for en reaktion i ligevægt. Deprotonering af His31 er en syre ligevægtsreaktion med en særlig Keq, der er kendt som syredissociationskonstanten, Ka: His31-H+ ⇌ His31 + H+. Den pKa er derefter relateret til Ka ved følgende: pKa = -log(Ka). Beregningen af forskellen i den frie energi mellem mutanten og wildtypen kan nu foretages ved hjælp af ligningen for den frie energi, definitionen af pKa, de observerede pKa-værdier og forholdet mellem naturlige logaritmer og logaritmer. I eksemplet med T4-lysozym gav denne fremgangsmåde et beregnet bidrag på ca. 3 kcal/mol til den samlede frie energi. En lignende fremgangsmåde kan anvendes med den anden deltager i saltbroen, f.eks. Asp70 i T4 lysozym-eksemplet, ved at overvåge dens forskydning i pKa efter mutation af His31.
Et forsigtighedsråd ved valg af det passende eksperiment omfatter saltbroens placering i proteinet. Miljøet spiller en stor rolle for interaktionen. Ved høje ioniske styrker kan saltbroen være fuldstændig maskeret, da der er tale om en elektrostatisk interaktion. His31-Asp70-saltbroen i T4 lysozym var begravet i proteinet. Entropi spiller en større rolle i overfladesaltbroer, hvor rester, der normalt har mulighed for at bevæge sig, er indsnævret af deres elektrostatiske interaktion og hydrogenbinding. Dette har vist sig at mindske entropien så meget, at interaktionens bidrag næsten udviskes. Overfladesaltbroer kan studeres på samme måde som begravede saltbroer ved hjælp af dobbelte mutantcyklusser og NMR-titreringer. Selv om der findes tilfælde, hvor begravede saltbroer bidrager til stabiliteten, findes der som i alle andre tilfælde undtagelser, og begravede saltbroer kan have en destabiliserende virkning. Også overfladesaltbroer kan under visse betingelser have en stabiliserende virkning. Den stabiliserende eller destabiliserende virkning skal vurderes fra sag til sag, og der kan kun fremsættes få generelle udsagn.