
Pterosiinien johdannaisten erilaistaminen ja karakterisoiminen pterosinin johdannaisilta, jotka ovat peräisin P. aquilinum
Pterosiinijohdannaiset eristettiin P. aquilinum -kasvin kokonaisista kasveista. Ahdekaunokin fytokemiallisten ainesosien tutkimiseksi vesiuute jaettiin peräkkäin etyyliasetaatilla (EtOAc) ja n-butanolilla (n-BuOH) (kuva 1). EtOAc-liukoisen fraktion toistuvalla kolonnikromatografialla eristettiin yhdeksän johdannaista, mukaan lukien (2R)-pterosiini B (1), pterosiini Z (2) ja (2S)-pterosiini P (3), (3R)-pterosiini D (4), (2S)-pterosiini A (5), (2S,3R)-pterosiini C (6), (2R,3R)-pterosiini C (7), (2R)-pterosidi B (8) ja pterosidi Z (9), joiden puhtausasteet olivat suurempia kuin 97 % (kuva. 2). N-BuOH-liukoisen fraktion toistuvalla kolonnikromatografisella erottelulla eristettiin kuusi johdannaista, mukaan lukien (2S)-pterosidi A (10), (2S)-pterosidi A2 (11), (2S,3R)-pterosidi C (12), (2R,3R)-pterosidi C (13), (3S)-pterosidi D (14) ja (2S)-pterosidi P (15), joiden puhtausasteet olivat suurempia kuin 97 %. Yhdisteiden rakenteet tunnistettiin analysoimalla spektritietoja, mukaan lukien MS, 1D- ja 2D-NMR (lisätiedot 1).

Pterosiiniyhdisteiden 1-15xxx rakenteet
Pterosiinijohdannaisten inhiboiva aktiivisuus BACE1:tä, AChE:tä vastaan, ja BChE
AD:n vastaisen potentiaalin arvioimiseksi kunkin pterosiiniyhdisteen inhiboiva aktiivisuus BACE1:tä ja koliiniesteraaseja vastaan arvioitiin vastaavilla in vitro -inhibitiomäärityksillä (taulukko 1). Kaikki testatut pterosiinijohdannaiset osoittivat pitoisuusriippuvaista inhiboivaa aktiivisuutta BACE1:tä vastaan IC50-arvojen (puoliksi suurin inhiboiva pitoisuus) vaihteluvälillä 9,74-94,4 μM, lukuun ottamatta (2S)-pterosiini A:ta ja (2S)-pterosidi P:tä, jotka olivat testatuilla pitoisuuksilla inaktiivisia verrattuna positiivisena kontrollina käytetyn kvertsetiinin IC50-arvoon, joka oli 18,8 μM. Voimakkaimpien inhibiittorien inhiboiva teho oli järjestyksessä (2R,3R)-pterosidi C, (3S)-pterosidi D, (2R)-pterosidi B, (2S,3R)-pterosiini C, (2R,3R)-pterosiini C, (2S,3R)-pterosidi C, (2S,3R)-pterosidi C ja (2R)-pterosiini B IC50-arvoilla, jotka olivat 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 ja 29,6 μM. Tämän jälkeen testasimme pterosiinijohdannaisten estopotentiaalia AChE:tä vastaan. Kaikki testatut yhdisteet osoittivat merkittäviä AChE:tä estäviä vaikutuksia, joiden IC50-arvot olivat välillä 2,55-110 μM, verrattuna positiivisena kontrollina käytetyn berberiinin IC50-arvoon AChE:tä vastaan, joka oli 0,39 μM. Pterosiiniyhdisteet, joilla oli voimakkain inhiboiva vaikutus AChE:tä vastaan, olivat (2R)-pterosidi B, (2R,3R)-pterosidi C, (2S,3R)-pterosidi C, (2S,3R)-pterosiini C ja (2R)-pterosiini B, joiden IC50-arvot olivat vastaavasti 2,55, 3,77, 9,17, 12,8 ja 16,2 μM. Lopuksi testasimme pterosiinijohdannaisten estokykyä BChE:tä vastaan. Kaikki testatut pterosiiniyhdisteet osoittivat inhiboivaa aktiivisuutta BChE:tä vastaan, ja IC50-arvot vaihtelivat välillä 5,29-119 μM, lukuun ottamatta (3R)-pterosiini D:tä, joka oli inaktiivinen testatuilla pitoisuuksilla verrattuna berberiinin IC50-arvoon BChE:tä vastaan, joka oli 3,32 μM. Pterosiiniyhdisteet, joilla oli voimakkain estävä vaikutus BChE:tä vastaan, olivat (2R,3R)-pterosidi C ja pterosidi Z, joiden IC50-arvot olivat 5,29 ja 5,31 μM.
Kokonaisuutena useimmilla testatuilla pterosiinijohdannaisilla oli merkittävää inhiboivaa vaikutusta BACE1:tä, AChE:tä ja BChE:tä vastaan samanaikaisesti. Ylimääräisen 2-hydroksimetyylitetrahydro-pyran-3,4,5-trioliryhmän läsnäolo kuten pterosidijohdannaisissa lisäsi merkittävästi inhiboivia vaikutuksia entsyymejä vastaan. Lisäksi ylimääräisen hydroksimetyyliryhmän läsnäolo (2R)-pterosiini B:n indanonirenkaan asemassa 2 kuten (2S)-pterosiini A:ssa tai metyyliryhmän läsnäolo kuten (3R)-pterosiini D:ssä ja pterosiini Z:ssä vähensi entsyymejä estäviä vaikutuksia. Erityisesti hydroksimetyyliryhmän läsnäolo indanonirenkaan positiossa 5, kuten (2S)-pterosiini P:ssä, vähensi inhiboivaa aktiivisuutta BACE1:tä vastaan.
Pterosiinijohdannaisten entsyymi-inhibition kineettiset parametrit
Yritimme selittää pterosiinijohdannaisten entsyymi-inhibitiotapaa tekemällä kineettisen analyysin edustaville inhibiittoreille BACE1:n ja koliiniesteraasien osalta (taulukko 2, lisätiedot 2). Alhainen Ki (inhibitiovakio) viittaa tiukempaan entsyymiin sitoutumiseen ja tehokkaampaan inhibiittoriin. Kaiken kaikkiaan yhdisteiden Ki-arvot korreloivat vastaavien IC50-arvojen kanssa. Yhdisteiden (2R,3R)-pterosidi C, (3S)-pterosidi D ja (2R,3R)-pterosiini C aiheuttama BACE1:n inhibitio oli sekatyyppistä, ja Ki-arvot olivat 12,6, 16,5 ja 27.6 µM, kun taas (2R)-pterosidi B:n, (2S,3R)-pterosiini C:n ja (2R)-pterosiini B:n aiheuttama inhibitio oli ei-kilpaileva, kun Ki-arvot olivat 23,1, 33,8 ja 38,3 µM. AChE:n esto (2R)-pterosidi B:llä, (2R,3R)-pterosidi C:llä, (2R)-pterosiini B:llä, (2S,3R)-pterosiini C:llä ja (3S)-pterosidi D:llä oli sekamuotoinen, ja sen Ki-arvot olivat 4.89, 8,13, 12,1, 16,3 ja 23,1 µM, kun taas (2R,3R)-pterosiini C oli ei-kilpaileva inhibiittori, jonka Kiarvo oli 29,6 µM. BChE:n esto (2R,3R)-pterosiini C:llä, (2R,3R)-pterosidi C:llä, (3S)-pterosidi D:llä ja (2R)-pterosidi B:llä oli sekamuotoista, ja niiden Ki-arvot olivat 4,77, 9,62, 19.7 ja 22,6 µM, kun taas (2S,3R)-pterosiini C ja (2R)-pterosiini B olivat ei-kilpailevia inhibiittoreita, joiden Kiarvot olivat 29,9 ja 53,5 µM. Näin ollen nämä tulokset viittasivat siihen, että tietyt pterosiinijohdannaiset saattavat olla tehokkaita BACE1-, AChE- ja BChE-estäjiä.
Molekyylien telakointisimulaatiot BACE1:lle, AChE:lle ja BChE:lle
BACE1:stä ja koliiniesteraaseista on saatavissa useita kiderakenteita. Valitsimme ihmisen PDB:t villityyppirakenteiden, yhteiskiteytettyjen ligandien ja rakenteiden resoluutioiden perusteella. Telakointia varten valittiin röntgenkiderakenteet BACE1:stä, joka on kompleksissa QUD:n kanssa (PDB-koodi: 2WJO, resoluutio: 2,5 Å)33 , AChE:stä, joka on kompleksissa E2020:n kanssa (PDB-koodi: 4EY7, resoluutio: 2,35 Å)34 , ja BChE:stä, joka on kompleksissa 3F9:n kanssa (PDB-koodi: 4TPK, resoluutio: 2,70 Å)35 . Aluksi QUD, E2020 ja 3F9 poimittiin kiderakenteista ja telakoitiin uudelleen BACE1:n, AChE:n ja BChE:n aktiivisiin keskuksiin. Tämän jälkeen (2R,3R)-pterosidi C, (3S)-pterosidi D, (2R,3R)-pterosiini C, (2R)-pterosidi B, (2S,3R)-pterosiini C ja (2R)-pterosiini B, joilla on tunnetut estomekanismit BACE1:tä, AChE:tä ja BChE:tä vastaan, telakoitiin niiden SAR-arvon määrittämiseksi. Taulukossa 3 on yhteenveto telakointituloksista. Valittujen sekatyyppisten ja ei-kilpailevien BACE1-, AChE- ja BChE-inhibiittoreiden SAR-arvot mahdollistivat pterosiinien suorittaman entsyymien uudenlaisen sitoutumiskohtiin kohdistuvan inhiboinnin arvioinnin.
E2020:n telakoitumismoodimme oli yhteneväinen aiemmin raportoidun kokeellisesti määritellyn sitoutumismoodin kanssa, joka on raportoitu rekombinantti-ihmisen AChE:llä (rhAChE) (Lisätiedot 3)34. E2020:n kide- ja telakoitujen konformaatioiden välinen neliöllinen keskihajonta (RMSD) oli 0,54 Å, mikä viittaa telakointiasetelmamme luotettavuuteen kokeellisen sitoutumistavan toistamisessa. Lisäksi E2020:n telakoitu tila johti samanlaiseen vuorovaikutukseen kuin rhAChE-E2020:n. Tutkimuksessamme vesimolekyylit poistettiin kiderakenteesta telakoinnin aikana, joten veden välittämiä vuorovaikutuksia ei analysoitu tässä tutkimuksessa. Vastaavasti QUD:n ja 3F9:n telakoituneet moodit olivat yhdenmukaisia BACE1 33:n ja BChE35:n osalta saatavilla olevien kokeellisten tietojen kanssa (lisätiedot 3). QUD:n ja 3F9:n kide- ja telakoitujen konformaatioiden väliset RMSD:t olivat 0,46 ja 0,60 Å. Lisäksi pterosiini-inhibiittorien sitoutumispaikat olivat sopusoinnussa aiemman telakointitutkimuksen kanssa, jossa oli mukana BACE1, AChE ja BChE38. Tutkimuksessa käytettiin kuitenkin Tetronarce californica AChE:tä (PDB-koodi: 1ACJ), joka sisältää hieman erilaisia jäännöslukuja kuin ihmisen AChE niiden sekvenssien vaihteluista johtuen.
BACE1:n telakointi
Inhibiittorityypin ja -aktiivisuuden perusteella (2R,3R)-pterosidi C ja (2R)-pterosidi B valittiin havainnollistamaan telakoituja moodeja sekatyyppisten ja ei-kilpailukykyisten BACE1:n inhibiittoreiden osalta vastaavasti. Kuvassa 3a, b esitetään (2R,3R)-pterosidi C:n ja (2R)-pterosidi B:n telakointimallit. Telakoitujen yhdisteiden vuorovaikutukset BACE1:n aktiivisen keskuksen sisällä on esitetty kuvassa 4.

Molekyylidockausmallit a sekatyyppiselle BACE1-inhibiittorille (2R,3R)-pterosidi C (sininen väri) ja b ei-kompetitiiviselle BACE1-inhibiittorille (2R)-pterosidi B (ruskea väri). Telakoidut asennot on asetettu QUD:n röntgenkiderakenteen päälle (keltainen väri) (PDB-koodi: 2WJO). BACE1, aktiivisen alueen jäännökset ja yhdisteet on esitetty nauhamalleilla, viivamalleilla ja sauvamalleilla. Pisteviivojen värit selittävät eri vuorovaikutustyypit: vetysidosvuorovaikutukset (vihreä), hydrofobiset vuorovaikutukset (vaaleanpunainen) ja π-sigma-vuorovaikutukset (violetti). BACE1 β-kohdan amyloidin esiasteproteiinia pilkkova entsyymi 1

Ligandin vuorovaikutuskaavio a QUD:n, b (2R,3R)-pterosidi C:n, c (3S)-pterosidi D:n, d (2R,3R)-pterosiini C:n, e (2R)-pterosidi B:n, f (2S,3R)-pterosiini C:n ja g (2R)-pterosiini B:n vuorovaikutuskaavio aktiivisessa keskuksessa BACE1:ssä. Katkoviivojen värit selittävät eri vuorovaikutustyypit: vetysidosvuorovaikutukset (vihreä), hydrofobiset vuorovaikutukset (vaaleanpunainen) ja π-sigma-vuorovaikutukset (violetti). BACE1 β-kohdan amyloidin esiasteproteiinia pilkkova entsyymi 1
QUD:n telakoidun poseerauksen sidosenergia (B.E.) oli -7,59 kcal/mol. Kuten kuvasta 4a näkyy, ligandin kinatsoliinirenkaan NH2-ryhmällä oli kaksi vetysidosta Asp32:n ja Asp228:n CO-ryhmien kanssa etäisyyksillä 1,86 ja 2,17 Å. Kolmas vetysidos havaittiin ligandin toisen NH-ryhmän ja Gly230:n CO-ryhmän välillä etäisyydellä 2,16 Å. Leu30, Tyr71, Phe108 ja Val332 välittivät hydrofobisia vuorovaikutuksia. Kuvissa 4b-d esitetään (2R,3R)-pterosidi C:n, (3S)-pterosidi D:n ja (2R,3R)-pterosiini C:n (sekatyyppiset BACE1-inhibiittorit) telakoidut asennot. Ne sijoittuivat sitoutumistaskuun, jota reunustavat Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 ja Thr231. Aktiivisuustasojensa perusteella (2R,3R)-pterosidi C:n (IC50 = 9,74 µM), (3S)-pterosidi D:n (IC50 = 10,7 µM) ja (2R,3R)-pterosiini C:n (IC50 = 26,2 µM) B.E. oli vastaavasti -7,27, -6,93 ja -4,84 kcal/mol. (2R,3R)-Pterosidi C:llä oli suurempi teho kuin (2R,3R)-Pterosiini C:llä, mikä johtui ylimääräisestä 2-hydroksimetyylitetrahydro-pyran-3,4,5-trioliryhmästä, joka muodosti kolme vetysidosta (kuva 4b). Hydroksimetyylin OH-ryhmä ja tetrahydropyraanitriolirenkaan 3-OH-ryhmä muodostivat kaksi vetysidosta Asp228:n CO-ryhmän kanssa etäisyyksillä 2,63 ja 2,12 Å. Lisäksi 3-OH-ryhmä osoitti vetysidoksen Thr231:n kanssa 1,89 Å:n etäisyydellä. (3S)-pterosidi D:n tapauksessa (kuva 4c) 2,2-dimetyyliryhmän läsnäolo indanonirenkaassa muutti hieman sitoutumisvuorovaikutuksia verrattuna (2R,3R)-pterosidi C:hen. Tetrahydropyraanitriolirenkaan 3-OH-ryhmä osoitti vain yhden vetysidoksen Asp228:n kanssa 1,93 Å:n päässä. Indanonirenkaan 3-OH-ryhmällä oli kuitenkin ylimääräinen vetysidos Ile126:n kanssa 2,34 Å:n etäisyydellä. Nämä vuorovaikutukset heikensivät hieman (3S)-pterosidi D:n aktiivisuutta verrattuna (2R,3R)-pterosidi C:hen. Kuvassa 4d esitetyt (2R,3R)-pterosiini C:n telakointivuorovaikutukset osoittavat, että indanonirenkaan 3-OH-ryhmä on sitoutunut Ser36:een ja Asn37:ään kahden vetysidoksen kautta 1,83 ja 2,07 Å:n etäisyydellä. Ala39, Val69, Trp76, Ile118 ja Arg128 olivat mukana hydrofobisissa vuorovaikutuksissa, kun taas Val69:llä oli π-sigma-vuorovaikutus.
(2R)-pterosidi B:n, (2S,3R)-pterosiini C:n ja (2R)-pterosiini B:n (ei-kilpailukykyiset BACE1:n estäjät) dokkaroidut poseeraukset on esitetty kuvissa 4e-g. Ne kiinnittyivät Asn37:n, Val69:n, Tyr71:n, Trp76:n, Lys107:n, Phe108:n ja Ile126:n rajaamaan onteloon. Aktiivisuusarvojensa mukaisesti (2R)-pterosidi B:n (IC50 = 18,0 µM), (2S,3R)-pterosiini C:n (IC50 = 23,1 µM) ja (2R)-pterosiini B:n (IC50 = 29,6 µM) B.E. oli vastaavasti -6,16, -5,07 ja -4,64 kcal/mol. Kuten kuvassa 4e näkyy, (2R)-pterosidi B osoitti suurempaa aktiivisuutta kuin (2S,3R)-pterosiini C ja (2R)-pterosiini B, mikä johtui ylimääräisen 2-hydroksimetyylitetrahydro-pyran-3,4,5-trioliryhmän läsnäolosta, joka osoitti neljää vetysidosvuorovaikutusta. Kaksi vetysidosta havaittiin tetrahydro-pyran-triolirenkaan 4-OH-ryhmän ja Asn37:n NH- ja CO-ryhmien välillä etäisyyksillä 2,48 ja 2,19 Å. Lisäksi 3- ja 5-OH-ryhmät osoittivat kaksi muuta vetysidosta Ile126:n ja Trp76:n kanssa etäisyyksillä 2,19 ja 2,46 Å. (2S,3R)-Pterosiini C osoitti hieman parempaa aktiivisuutta kuin (2R)-Pterosiini B, mikä johtui ylimääräisen OH-ryhmän läsnäolosta indanonirenkaan positiossa 3, joka muodosti vetysidoksen Lys107:n kanssa 2,12 Å:n etäisyydelle (kuva 4f). Muut vuorovaikutukset olivat samanlaisia kuin (2R)-pterosiini B:llä. Kuten kuvassa 4g näkyy, (2R)-pterosiini B:llä oli hydrofobisia vuorovaikutuksia Val69:n, Tyr76:n ja Phe108:n kanssa.
AChE:n telakointi
(2R)-Pterosidi B ja (2R,3R)-pterosiini C valittiin edustajiksi osoittamaan sekatyyppisten ja ei-kilpailukykyisten AChE:n estäjien telakointitapoja niiden aktiivisuuden ja AChE:n eston tyypin vuoksi. Kuvissa 5a, b on esitetty (2R)-pterosidi B:n ja (2R,3R)-pterosiini C:n telakointimallit. Telakoitujen yhdisteiden vuorovaikutukset AChE:n aktiivisen keskuksen sisällä on esitetty kuvassa 6.

Molekulaariset telakointimallit a sekamuotoiselle AChE:n estäjälle (2R)-pterosidi B (sininen väri) ja b ei-kompetitiiviselle AChE:n estäjälle (2R,3R)-pterosiini C (ruskea väri). Telakoituneet asennot on sijoitettu E2020:n röntgenkiderakenteen päälle (keltainen väri) (PDB-koodi: 4EY7). AChE, aktiivisen alueen jäännökset ja yhdisteet on esitetty nauhamalleilla, viivamalleilla ja tikkumalleilla. Pisteviivojen värit selittävät eri vuorovaikutustyypit: vetysidokset (vihreä) ja hydrofobiset vuorovaikutukset (vaaleanpunainen). AChE asetyylikoliiniesteraasi

Ligandin vuorovaikutuskaavio a E2020, b (2R)-pterosidi B, c (2R,3R)-pterosidi C, d (2S,3R)-pterosiini C, e (2R)-pterosiini B, f (3S)-pterosidi D ja g (2R,3R)-pterosiini C AChE:n aktiivisessa keskuksessa. Katkoviivojen värit selittävät eri vuorovaikutustyypit: vetysidosvuorovaikutukset (vihreä), hydrofobiset vuorovaikutukset (vaaleanpunainen) ja π-sigma-vuorovaikutukset (violetti). AChE asetyylikoliiniesteraasi
E2020:n telakoitu poseeraus osoitti B.E.:n olevan -10,28 kcal/mol. Kuten kuvasta 6a käy ilmi, indanonirenkaan CO-ryhmä muodosti vetysidoksen Phe295:n NH-ryhmän kanssa 1,70 Å:n etäisyydellä. Trp286 ja Tyr341 osallistuivat π-sigma-vuorovaikutuksiin, kun taas Trp86, Trp286, Tyr337, Phe338 ja Tyr341 välittivät hydrofobisia vuorovaikutuksia. Kuvissa 6b-f on esitetty (2R)-pterosidi B:n, (2R,3R)-pterosidi C:n, (2S,3R)-pterosiini C:n, (2R)-pterosiini B:n ja (3S)-pterosidi D:n (sekatyyppiset AChE:n estäjät) telakoidut asennot. Ne sijoittuivat Tyr72:n, Asp74:n, Trp86:n, Asn87:n, Tyr124:n, Trp286:n, Phe295:n, Phe297:n, Tyr337:n, Phe338:n ja Tyr341:n ympäröimään aktiiviseen kohtaan. Aktiivisuusarvojensa mukaisesti (2R)-pterosidi B (IC50 = 2,55 µM), (2R,3R)-pterosidi C (IC50 = 3,77 µM), (2S,3R)-pterosiini C (IC50 = 12.8 µM), (2R)-pterosiini B (IC50 = 16,2 µM) ja (3S)-pterosidi D (IC50 = 27,4 µM) B.E. oli vastaavasti -7,90, -7,49, -6,03, -5,76 ja -4,91 kcal/mol. (2R)-Pterosidi B osoitti suurempaa tehoa kuin (2S,3R)-Pterosiini C ja (2R)-Pterosiini B, mikä johtui ylimääräisen 2-hydroksimetyylitetrahydro-pyran-3,4,5-trioliryhmän läsnäolosta, joka synnytti kolme vetysidevuorovaikutusta (kuva 6b). Hydroksimetyylin OH-ryhmä ja tetrahydropyraanitriolirenkaan 3-OH-ryhmä muodostivat vetysidoksia Asn87:n CO-ryhmän kanssa 2,23 ja 2,28 Å:n etäisyydellä. Lisäksi 4-OH-ryhmä muodosti vetysidoksen Trp86:n CO-ryhmän kanssa 2,13 Å:n etäisyydellä. (2R,3R)-pterosidi C:n (kuva 6c) tapauksessa indanonirenkaan asemassa 2 olevalla metyyliryhmällä ei ollut hydrofobista vuorovaikutusta Tyr341:n kanssa, ja sen aktiivisuus oli näin ollen verrattain alhaisempi kuin (2R)-pterosidi B:llä. Tyr341 säilytti kuitenkin hydrofobisen vuorovaikutuksen indanonirenkaan muun osan kanssa, kuten (2R)-pterosidi B:n tapauksessa. (2S,3R)-pterosiini C:n korkeampi aktiivisuus kuin (2R)-pterosiini B:n johtui siitä, että indanonirenkaan positiossa 3 oli ylimääräinen OH-ryhmä, joka muodosti vetysidoksen Phe295:n kanssa etäisyydelle 1,82 Å (kuva 6d). Muut vuorovaikutukset olivat verrattavissa (2R)-pterosiini B:n vuorovaikutuksiin. Kuten kuvasta 6e nähdään, indanonirenkaan asemassa 6 olevan hydroksietyyliryhmän OH-ryhmä muodosti vetysidoksen Tyr124:n kanssa 2,48 Å:n etäisyydelle. Tyr124, Trp286, Tyr337, Phe338 ja Tyr341 osallistuivat hydrofobisiin vuorovaikutuksiin. Verrattuna (2R)-pterosidi B:hen ja (2R,3R)-pterosidi C:hen (3S)-pterosidi D:llä oli erilaiset sitoutumisvuorovaikutukset, jotka johtuivat 2,2-dimetyyliryhmän läsnäolosta indanonirenkaassa (kuva 6f). 2,2-dimetyyliryhmä vaikutti merkittävästi (3S)-pterosidi D:n erilaiseen telakoituneeseen asentoon. Tetrahydropyraanitriolirenkaan 5-OH-ryhmä muodosti vetysidoksen Asp74:n kanssa 1,96 Å:n etäisyydelle. Indanonirenkaan CO-ryhmä muodosti vetysidoksen Tyr72:n kanssa etäisyydellä 2,97 Å. Nämä vuorovaikutukset selittivät (3S)-pterosidi D:n alhaisen aktiivisuuden.
(2R,3R)-pterosiini C:n (ei-kilpailukykyinen AChE:n estäjä) telakoitunut poseeraus on esitetty kuvassa 6g. (2R,3R)-Pterosiini C:n (IC50 = 23,2 µM) B.E. oli -5,01 kcal/mol. (2R,3R)-pterosiini C:n sitoutumistasku käsitti Tyr72:n, Trp286:n, Ser293:n, Phe297:n ja Tyr341:n, joissa oli kaksi vetysidosvuorovaikutusta. Toinen vetysidoksista muodostui indanonirenkaan 3-OH-ryhmän ja Ser293:n CO-ryhmän välille 2,03 Å:n etäisyydelle. Toinen vetysidos havaittiin indanonirenkaan asemassa 6 olevan hydroksietyyliryhmän OH-ryhmän ja Tyr72:n välillä 1,94 Å:n etäisyydellä. Tyr72:n, Trp286:n, Phe297:n ja Tyr341:n kaltaiset jäännökset osallistuivat hydrofobisiin vuorovaikutuksiin.
BChE:n telakointi
Aktiivisuustasot ja BChE:n inhibitiotyyppi huomioon ottaen (2R,3R)-pterosidi C ja (2S,3R)-pterosiini C valittiin havainnollistamaan sekatyyppisten sekatyyppisten ja ei-kilpailukykyisten BChE:n inhibiittoreiden telakoituja moodeja. Kuvissa 7a, b esitetään (2R,3R)-pterosidi C:n ja (2S,3R)-pterosiini C:n telakointimallit. Telakoitujen yhdisteiden vuorovaikutukset BChE:n aktiivisen keskuksen sisällä on esitetty kuvassa 8.
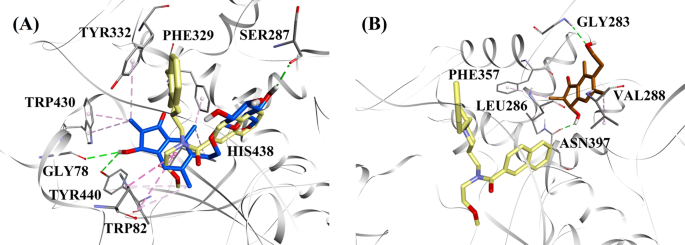
Molekulaariset telakointimallit a sekatyyppiselle BChE:n estäjälle (2R,3R)-pterosiini C (sininen väri) ja b ei-kilpailukykyiselle BChE:n estäjälle (2S,3R)-pterosiini C (ruskea väri). Telakoituneet asennot on sijoitettu 3F9:n röntgenkiderakenteen päälle (keltainen väri) (PDB-koodi: 4TPK). BChE, aktiivisen alueen jäännökset ja yhdisteet on esitetty nauhamalleilla, viivamalleilla ja sauvamalleilla. Pisteviivojen värit selittävät eri vuorovaikutustyypit: vetysidokset (vihreä) ja hydrofobiset vuorovaikutukset (vaaleanpunainen). BChE butyylikoliiniesteraasi
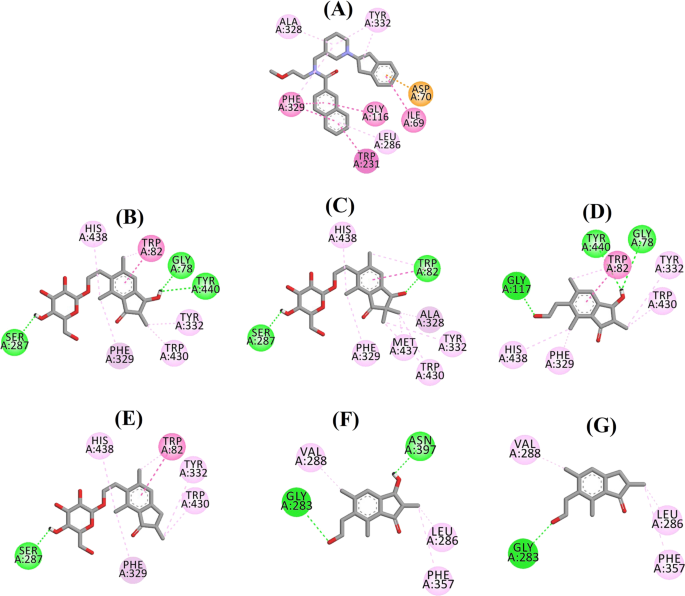
Ligandin vuorovaikutuskaavio a 3F9:n, b (2R,3R)-pterosidi C:n, c (3S)-pterosidi D:n, d (2R,3R)-pteerosiini C:n, e (2R)-pteerosidi B:n, f (2S,3R)-pteerosiini C:n ja g (2R)-pteerosiini B:n vuorovaikutuskaavio BChE:n aktiivisessa paikassa. Katkoviivojen värit selittävät eri vuorovaikutustyypit: vetysidosvuorovaikutukset (vihreä), hydrofobiset vuorovaikutukset (vaaleanpunainen) ja π-anionien vuorovaikutukset (kultainen). BChE butyylikoliiniesteraasi
3F9:n telakoidun asennon B.E. oli -8,49 kcal/mol. Kuten kuvassa 8a on esitetty, hydrofobiset vuorovaikutukset olivat pääasiassa vastuussa ligandin sitoutumisesta. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 ja Tyr332 vastasivat hydrofobisista vuorovaikutuksista, kun taas Asp70 osoitti π-anioni vuorovaikutusta. Kuvissa 8b-e on esitetty (2R,3R)-pterosidi C:n, (3S)-pterosidi D:n, (2R,3R)-pterosiini C:n ja (2R)-pterosidi B:n (sekatyyppiset BChE:n estäjät) telakoidut asennot. Niiden sitoutumistasku koostui Gly78:sta, Trp82:sta, Gly117:stä, Ser287:sta, Ala328:sta, Phe329:stä, Tyr332:sta, Trp430:sta, Met437:stä, His438:sta ja Tyr440:stä. Aktiivisuustasojensa mukaisesti (2R,3R)-pterosidi C (IC50 = 5,29 µM), (3S)-pterosidi D (IC50 = 19,3 µM), (2R,3R)-pterosiini C (IC50 = 20,3 µM) ja (2R)-pterosidi B (IC50 = 62,0 µM) osoittivat, että niiden B.E. oli vastaavasti -7,23, -6,59, -6,52 ja -4,38 kcal/mol. Kuten kuvasta 8b käy ilmi, (2R,3R)-pterosidi C:n indanonirenkaan 3-OH-ryhmässä oli kaksi vetysidosta Gly78:n ja Tyr440:n kanssa etäisyyksillä 2,87 ja 2,89 Å. (3S)-pterosidi D:n tapauksessa indanonirenkaan 2,2-dimetyyliryhmä vaikutti 3-OH-ryhmän sitoutumisvuorovaikutuksiin (kuva 8c). 3-OH-ryhmä muodosti vain yhden vetysidoksen Trp82:n kanssa 2,94 Å:n etäisyydelle, mikä johti (3S)-pterosidi D:n verrattain alhaisempaan aktiivisuuteen kuin (2R,3R)-pterosidi C:n. Kuten kuvasta 8d käy ilmi, (2R,3R)-pterosiini C ei pystynyt muodostamaan vetysidosta Ser287:n kanssa, koska siinä ei ollut 2-hydroksimetyylitetrahydropyraani-3,4,5-trioliryhmää. Näin ollen sen aktiivisuus oli alhaisempi kuin (2R,3R)-pterosidi C:n. OH-ryhmän puuttuminen indanonirenkaan positiosta 3 osoittautui syyksi (2R)-pterosidi B:n erittäin alhaiselle aktiivisuudelle (kuva. 8e), joka ei kyennyt muodostamaan vetysidoksia Gly78:n ja Tyr440:n kanssa samalla tavalla kuin (2R,3R)-pterosidi C.
(2S,3R)-pterosiini C:n ja (2R)-pterosiini B:n (ei-kilpailukykyiset BChE:n estäjät) telakoidut poseeraukset on esitetty kuvissa 8f, g. Nämä telakoituneet positiot olivat Gly283:n, Leu286:n, Val288:n, Phe357:n ja Asn397:n ympäröimässä ontelossa. Aktiivisuustasojen mukaan (2S,3R)-pterosiini C:n (IC50 = 44,3 µM) ja (2R)-pterosiini B:n (IC50 = 48,1 µM) B.E. oli vastaavasti -5,40 ja -5,06 kcal/mol. (2S,3 R)-Pterosiini C oli tehokkaampi kuin (2R)-pterosiini B, koska indanonirenkaan asemassa 3 oli ylimääräinen OH-ryhmä, joka muodosti vetysidoksen Asn397:n CO-ryhmän kanssa 2,05 Å:n etäisyydelle (kuva 8f). Muiden vuorovaikutusten havaittiin olevan samankaltaisia kuin (2R)-pterosiini B:ssä. Kuten kuvasta 8g nähdään, indanonirenkaan positiossa 6 olevan hydroksietyyliryhmän OH-ryhmä muodosti vetysidoksen Gly283:n kanssa 2,40 Å:n etäisyydelle. Jäännökset Leu286, Val288 ja Phe357 osallistuivat hydrofobisiin vuorovaikutuksiin.
Sekatyyppiset inhibiittorit sitoutuvat sekä vapaaseen entsyymiin että entsyymi-substraatti-kompleksiin, mikä osoittaa, että nämä yhdisteet voivat sitoutua kunkin vastaavan entsyymin katalyyttiseen kohtaan. Ei-kompetitiiviset inhibiittorit sitoutuvat vapaan entsyymin tai entsyymi-substraatti-kompleksin allosteriseen kohtaan. Tuoreessa tutkimuksessa esitettiin, että kilpailevat, sekatyyppiset ja ei-kilpailevat inhibiittorit miehittävät eri paikkoja BACE1:n, AChE:n ja BChE:n38 sitoutumistaskuissa. Pterosiinijohdannaisten inhibitiomekanismin arvioimiseksi tehdyssä telakoinnissa yhdisteiden sitoutumispaikat määriteltiin niiden inhibitiotyypin mukaan. Telakointitulokset osoittivat, että BACE1:n, AChE:n ja BChE:n sekatyyppisten ja ei-kilpailevien inhibiittoreiden sitoutumiskohteet ovat osittain päällekkäin kussakin vastaavassa aktiivisessa kohdassa, ja ne olivat yhdenmukaisia aikaisemman raportin kanssa38.
BBBB:n läpäisevyys
PAMPA-BBB, joka on in vitro -menetelmällä tehty keinotekoinen BBB:n kalvon läpäisevyystesti, on yksi luotettavimmista fysikokemiallis-kemiallisista seulontatutkimuslaitteistosta, jota käytetään keskushermostoon kohdistuvien lääkkeiden varhaisessa vaiheessa tapahtuvassa keksimisessä40. PAMPA-BBB-järjestelmä mallintaa kemikaalien transsellulaarista passiivista diffuusiota BBB:n läpi ja mittaa puhtaasti passiivisia kuljetusmekanismeja keinotekoisen lipidikalvon kautta efektiivisen läpäisevyyden (Pe, cm/s) perusteella. BBB-läpäisevyyden ennustamista varten laaditun mallin perusteella yhdisteet luokiteltiin seuraavasti: i) ”CNS+” (ennustettu korkea BBB-läpäisevyys); Pe (10-6 cm/s) > 4,00, ii) ”CNS-” (ennustettu matala BBB-läpäisevyys); Pe (10-6 cm/s) < 2,00 ja iii) ”CNS+/-” (BBB-läpäisevyys epävarmaa); Pe (10-6 cm/s) 4,00-2,00. Näin ollen (2R)-pterosiini B:llä, (2S)-pterosiini P:llä ja (2S)-pterosiini A:lla oli korkea BBB-permeaatio, kun Pe-arvot olivat vastaavasti 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s ja 6,26 × 10-6 cm/s (taulukko 4, lisätiedot 4). (2R)-pterosiini B:n Pe-arvo oli 1,7-kertainen verrattuna positiivisena kontrollina käytetyn keskushermostolääkkeen verapamiilin Pe-arvoon (Pe = 34,6 × 10-6 cm/s). (2S,3R)-Pterosiini C ja (2R,3R)-pterosiini C osoittivat epävarmaa BBB-permeaatiota Pe-arvoilla 2,34 ja 1,98. (2R,3R)-Pterosidi C:llä, (3S)-pterosidi D:llä ja (2R)-pterosidi B:llä, joilla oli testatuista pterosiinijohdannaisista voimakkaimmat BACE1- ja koliiniesteraasi-inhibiittorit, oli hyvin alhainen BBB-läpäisevyys. 2-hydroksimetyylitetrahydro-pyran-3,4,5-trioliryhmän esiintyminen pterosidijohdannaisissa johti BBB-läpäisevyyden huomattavaan vähenemiseen. Verrattuna (2R)-pterosiini B:hen indanonirenkaan OH-ryhmän läsnäolo indanonirenkaan asennossa 3 kuten pterosiini C:ssä, hydroksimetyyliryhmän läsnäolo indanonirenkaan asennossa 2 kuten (2S)-pterosiini A:ssa tai hydroksimetyyliryhmän läsnäolo indanonirenkaan asennossa 5 kuten (2S)-pterosiini P:ssä alensi myös merkittävästi BBB:n läpäisevyyttä. Kun otetaan huomioon poikkeuksellisen korkea BBB-läpäisevyys ja BACE1:n, AChE:n ja BChE:n merkittävä inhibitio, (2R)-pterosiini B:llä voi olla potentiaalia osoittaa vahvaa anti-AD-aktiivisuutta.
(2R)-pterosiini B:n ja (2R,3R)-pterosidi C:n vaikutus Aβ-peptidien erittymiseen hermosoluista
Tutkittaessa pterosiini B:n ja (2R,3R)-pterosidi C:n toimintaa
,3R)-pterosidi C:n vaikutusta Aβ:n erittymisen vähentämisessä hermosoluista, käytimme hiiren neuroblastooma-solulinjaa, joka yliekspressoi stabiilisti ihmisen APPswe:tä. Solulinja on AD:n solumalli, jolle on ominaista Aβ40:n ja Aβ42:n liiallinen eritys. Myrkylliset amyloidi-oligomeerit muodostuvat Aβ-peptidin kahdesta eri pituisesta isomuodosta. Aβ40 on aivojen runsain Aβ-isomuoto41, kun taas Aβ42 lisääntyy merkittävästi tietyissä Alzheimerin taudin muodoissa42. Aβ40:n sandwich-ELISA osoitti, että (2R)-pterosiini B vähensi merkittävästi neuroblastoomasoluista väliaineeseen erittyvän Aβ40-peptidin määrää jopa 50 prosentilla 500 μM:n annoksella (P < 0,01) (kuva 9a). Vastaavasti Aβ42-peptidin eritys neuroblastoomasoluista väheni merkittävästi, kun läsnä oli 500 μM (2R,3R)-pterosidi C:tä (P < 0,05) (Kuva 9b). Yhteenvetona voidaan todeta, että (2R)-pterosiini B ja (2R,3R)-pterosidi C vähensivät merkittävästi Aβ-peptidien eritystä neuroblastoomasoluista 500 μM:n pitoisuudessa.

a (2R)-pterosiini B:n vaikutus Aβ-peptidien eritykseen. APPswe-erittäviä neuroblastoomasoluja käsiteltiin 12, 60, 250 tai 500 µM (2R)-pterosiini B:llä 24 tunnin ajan, ja konditionoitu media kerättiin proteaasi-inhibiittorin läsnä ollessa. Positiivisena kontrollina käytettiin β-SI:tä (10 µM). Negatiiviseen kontrolliin kuuluivat solut, joita viljeltiin ilman testiyhdisteitä. Konditionoidussa mediassa erittyvän Aβ40:n ja Aβ42:n kvantitatiivinen analyysi tehtiin sandwich-ELISA:lla. Esitetään kolmen riippumattoman kokeen keskiarvot ± SD. Erittyvät Aβ40-peptidit vähenivät merkittävästi 500 µM (2R)-pterosiini B:n läsnäollessa. CON-negatiivinen kontrolli, β-SI β-sekretaasi-inhibiittori III, Aβ β-amyloidi. *P < 0,05, **P < 0,01. b (2R,3R)-pterosidi C:n vaikutus Aβ-peptidien eritykseen. APPswe-erittäviä neuroblastoomasoluja käsiteltiin 12, 60, 125, 250 tai 500 µM (2R,3R)-pterosidi C:llä 24 tunnin ajan, ja konditionoitu media kerättiin proteaasinestäjän läsnä ollessa. Positiivisena kontrollina käytettiin β-SI:tä (10 µM). Negatiiviseen kontrolliin kuuluivat solut, joita viljeltiin ilman testiyhdisteitä. Konditionoidussa väliaineessa erittyvän Aβ40:n ja Aβ42:n kvantitatiivinen analyysi tehtiin sandwich-ELISA-menetelmällä. Esitetään kolmen riippumattoman kokeen keskiarvot ± SD. Erittyvä Aβ42-peptidi väheni merkittävästi 500 µM (2R,3R)-pterosidi C:n läsnäollessa. CON-negatiivinen kontrolli, β-SI β-sekretaasi-inhibiittori III, Aβ β-amyloidi. *P < 0.05
Pterosiinijohdannaisten sytotoksisuus MTT-määrityksen perusteella43
Kokonaisuutena pterosiinijohdannaiset osoittivat mitättömän pientä sytotoksisuutta erilaisia normaaleja ja syöpäsolulinjoja, kuten SH-SY5Y-ihmisen neuroblastoomaa, C6-rotan gliasoluja, NIH3T3-hiiren alkiofibroblasteja ja B16F10-hiiren melanoomaa, vastaan, ja niiden LD50-arvot olivat suuremmat kuin 0,0.5 mM (lisätiedot 5). Erityisesti (2R)-pterosiini B, (3R)-pterosiini D, (2S)-pterosiini P, (2S)-pterosidi A, (2R)-pterosidi B ja (2R,3R)-pterosidi C eivät osoittaneet sytotoksisuutta testattuja solulinjoja vastaan, joiden LD50-arvot olivat yli 5 mM. Mielenkiintoista on, että useat pterosiinit osoittivat suhteellisia antiproliferatiivisia vaikutuksia SH-SY5Y-neuronisoluihin verrattuna mesenkymaalista alkuperää oleviin C6-gliasoluihin. Nämä tulokset olivat yhdenmukaisia aikaisemman raportin kanssa, jonka mukaan pterosiinijohdannaiset eivät ole ihmiselle myrkyllisiä26.