

Suolasillan osuutta proteiinin kokonaisstabiliteettiin taittuneeseen tilaan voidaan arvioida mutaatiotutkimuksista ja ydinmagneettiresonanssitekniikoilla kerätyn termodynaamisen tiedon avulla. Käyttämällä mutatoitua pseudo-wild-tyyppistä proteiinia, joka on erityisesti mutatoitu estämään saostuminen korkeassa pH:ssa, voidaan määrittää suolasillan osuus laskostuneen proteiinitilan kokonaisvapaaseen energiaan suorittamalla pistemutaatio, joka muuttaa ja näin ollen rikkoo suolasillan. Esimerkiksi T4-lyysotsyymissä tunnistettiin suolasilta jäännös 70:ssä olevan asparagiinihapon (Asp) ja jäännös 31:ssä olevan histidiinin (His) välillä (kuva 3). Paikkaohjattu mutageneesi asparagiinilla (Asn) (kuva 4) tehtiin ja saatiin kolme uutta mutanttia: Asp70Asn His31 (mutantti 1), Asp70 His31Asn (mutantti 2) ja Asp70Asn His31Asn (kaksoismutantti).

Kun mutantit on määritetty, voidaan käyttää kahta menetelmää suolasiltaan liittyvän vapaan energian laskemiseksi. Toisessa menetelmässä tarkkaillaan villityypin proteiinin sulamislämpötilaa verrattuna kolmen mutantin sulamislämpötilaan. Denaturoitumista voidaan seurata sirkulaaridikroismin muutoksen avulla. Sulamislämpötilan aleneminen osoittaa stabiilisuuden vähenemistä. Tämä kvantifioidaan Becktelin ja Schellmanin kuvaamalla menetelmällä, jossa kahden vapaan energian ero lasketaan ΔTΔS:n avulla. Tähän laskutoimitukseen liittyy joitakin ongelmia, ja sitä voidaan käyttää vain erittäin tarkkojen tietojen perusteella. T4-lyserotsyymin esimerkissä pseudo-wild-tyypin ΔS oli aiemmin raportoitu pH:ssa 5,5, joten 11 °C:n keskilämpötilaero tässä pH:ssa kerrottuna raportoidulla ΔS:llä, joka on 360 cal/(mol-K) (1,5 kJ/(mol-K)), saadaan vapaan energian muutokseksi noin -4 kcal/mol (-17 kJ/mol). Tämä arvo vastaa sitä vapaan energian määrää, jonka suolasilta vaikuttaa proteiinin stabiilisuuteen.
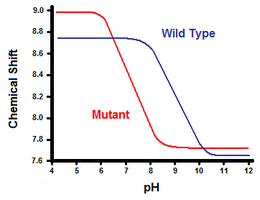
Toisessa menetelmässä hyödynnetään ydinmagneettista resonanssispektroskopiaa suolasillan vapaan energian laskemiseen. Suoritetaan titraus ja samalla rekisteröidään karboksylaatti- tai ammoniumryhmän vieressä olevan hiilen protoneja vastaava kemiallinen siirtymä. Titrauskäyrän keskipiste vastaa pKa:ta eli pH:ta, jossa protonoituneiden ja deprotonoituneiden molekyylien suhde on 1:1. Jatkamalla T4-lyserotsyymin esimerkkiä saadaan titrauskäyrä havaitsemalla histidiini 31:n C2-protonin siirtymä (kuva 5). Kuvassa 5 esitetään titrauskäyrän siirtymä villityypin ja sellaisen mutantin välillä, jossa Asp70 on Asn. Suolasilta muodostuu deprotonoituneen Asp70:n ja protonoituneen His31:n välille. Tämä vuorovaikutus aiheuttaa His31:n pKa:ssa havaitun muutoksen. Epäselvässä villityypin proteiinissa, jossa suolasilta puuttuu, His31:n pKa:n ilmoitetaan olevan 6,8 kohtalaisen ionivahvassa H20-puskurissa. Kuvassa 5 esitetään villityypin pKa:ksi 9,05. Tätä pKa:n eroa tukee His31:n vuorovaikutus Asp70:n kanssa. Säilyttääkseen suolasillan His31 pyrkii pitämään protoninsa mahdollisimman pitkään. Kun suolasilta katkeaa, kuten mutantissa D70N, pKa siirtyy takaisin arvoon 6,9, joka on paljon lähempänä His31:n arvoa avautumattomassa tilassa.
PKa:n ero voidaan kvantifioida siten, että se kuvastaa suolasillan osuutta vapaaseen energiaan. Käyttämällä Gibbsin vapaaenergiaa: ΔG = -RT ln(Keq), missä R on yleinen kaasuvakio, T on lämpötila kelvineinä ja Keq on tasapainossa olevan reaktion tasapainovakio. His31:n deprotonoituminen on hapon tasapainoreaktio, jolla on erityinen Keq, jota kutsutaan hapon dissosiaatiovakioksi, Ka: His31-H+ ⇌ His31 + H+. Tällöin pKa liittyy Ka:han seuraavasti: pKa = -log(Ka). Mutantin ja villityypin vapaan energian eron laskeminen voidaan nyt tehdä käyttämällä vapaan energian yhtälöä, pKa:n määritelmää, havaittuja pKa:n arvoja sekä luonnollisen logaritmin ja logaritmin välistä suhdetta. T4-lyysotsyymin esimerkissä tämä lähestymistapa antoi laskennallisen noin 3 kcal/mol:n osuuden kokonaisvapaaseen energiaan. Samanlainen lähestymistapa voidaan käyttää suolasillan toiselle osallistujalle, kuten Asp70:lle T4-lysotsyymiesimerkissä, seuraamalla sen pKa:n muutosta His31:n mutaation jälkeen.
Varoituksen sana sopivaa koetta valittaessa liittyy suolasillan sijaintiin proteiinissa. Ympäristöllä on suuri merkitys vuorovaikutuksessa. Suurilla ionivahvuuksilla suolasilta voi peittyä kokonaan, koska kyseessä on sähköstaattinen vuorovaikutus. T4-lyysotsyymin His31-Asp70-suolasilta oli hautautunut proteiinin sisään. Entropialla on suurempi merkitys pintasuolasilloissa, joissa jäännökset, joilla on normaalisti mahdollisuus liikkua, ovat rajoittuneet sähköstaattisen vuorovaikutuksen ja vetysidoksen vuoksi. Tämän on osoitettu vähentävän entropiaa niin paljon, että vuorovaikutuksen vaikutus lähes häviää. Pintasuolasiltoja voidaan tutkia samalla tavalla kuin upotettuja suolasiltoja käyttämällä kaksoismutanttisyklejä ja NMR-titrauksia. Vaikka on olemassa tapauksia, joissa hautautuneet suolasillat edistävät stabiilisuutta, kuten kaikesta muustakin, poikkeuksia on olemassa, ja hautautuneilla suolasilloilla voi olla destabiloiva vaikutus. Myös pintasuolasilloilla voi tietyissä olosuhteissa olla stabiloiva vaikutus. Stabiloiva tai destabiloiva vaikutus on arvioitava tapauskohtaisesti, eikä yleisiä lausuntoja voida antaa.