
Introduction
La mémoire autobiographique est souvent décrite en termes de deux types de mémoire à long terme, la mémoire sémantique (connaissances sur le soi) et la mémoire épisodique (connaissances spécifiques à un événement liées à des expériences personnelles passées) (Tulving, 2002). La composante de la mémoire épisodique est considérée comme la caractéristique déterminante de la récupération de la mémoire autobiographique, car elle permet de se souvenir d’événements passés de manière très détaillée (Conway, 2001 ; Rubin, 2005). Lors de la remémoration, les processus de mémoire épisodique reconstruisent activement une expérience autobiographique en associant ensemble différents détails expérientiels, y compris les éléments perceptifs et conceptuels (Bartlett, 1932 ; Schacter et Addis, 2007 ; Schacter et al., 2011 ; Sheldon et Levine, 2016). Ici, nous proposons que cette caractéristique reconstructive de la mémoire épisodique permet différentes formes de souvenir autobiographique en construisant des représentations de mémoire avec différentes combinaisons de détails. Plus précisément, nous suggérons que les événements autobiographiques peuvent être représentés et mémorisés en tant qu’expériences conceptuelles ou perceptuelles et que ces formes : (a) s’appuient sur des mécanismes neuronaux différents ; et (b) contribuent à différentes fonctions de la mémoire, en particulier lorsque la mémoire est utilisée pour résoudre un problème actuel ou diriger une action future (par exemple, Vandermorris et al., 2013 ; Madore et al., 2016 ; Schacter et al., 2017 ; Mar et Spreng, 2018).
Dans les sections qui suivent, nous développons ces deux points en explorant la nature et les raisons d’une division perceptuelle/conceptuelle au sein de la mémoire autobiographique épisodique. Nous examinons d’abord comment ces différentes représentations autobiographiques épisodiques (une représentation conceptuelle et une représentation perceptuelle) émergent des théories de l’organisation de la mémoire autobiographique. Nous décrivons ensuite les résultats de la neuro-imagerie qui suggèrent que ces formes de mémoire correspondent à des systèmes de traitement de l’information dissociables dans le cerveau. Nous passons également en revue les travaux qui montrent comment l’hippocampe antérieur et postérieur facilite l’activité au sein de ces systèmes de traitement à grande échelle. Enfin, inspirés par la recherche sur la façon dont la mémoire autobiographique épisodique sert une variété de fonctions non mémorielles, y compris l’orientation des décisions et des comportements futurs (par exemple, Pillemer, 2003 ; Alea et Bluck, 2007), nous discutons des scénarios de prise de décision qui bénéficient de l’adoption d’une forme perceptive par rapport à une forme conceptuelle de mémorisation. Ici, nous notons également comment les distinctions dans la mémorisation peuvent s’étendre à d’autres fonctions autobiographiques non-directives (auto et sociales).
Théories de l’accès et de l’organisation de la mémoire autobiographique
Lors de la récupération des expériences autobiographiques, la mémoire épisodique soutient la capacité de se rappeler richement une expérience telle qu’elle s’est produite pendant un temps et un lieu spécifiques (Tulving, 2002 ; Szpunar et McDermott, 2009). Pendant la récupération, ces processus de mémoire épisodique construisent une représentation détaillée de la mémoire en associant différents types d’informations sur l’événement traitées par des systèmes de composants disparates (par exemple, détails visuels, détails auditifs, informations conceptualisées ; Greenberg et Rubin, 2003 ; Rubin, 2005 ; Moscovitch et al., 2016). Cette caractéristique constructive de la mémoire épisodique signifie que de multiples types de représentations de la mémoire autobiographique peuvent être formés en engageant différentes combinaisons – et pondérations relatives – des processus composants (Rubin, 2006 ; St. Jacques et al., 2011 ; Cabeza et Moscovitch, 2013 ; Moscovitch et al., 2016).
Une théorie de l’organisation des connaissances autobiographiques suggère qu’une distinction dans la façon dont les représentations de la mémoire autobiographique sont formées est en tant qu’événements épisodiques principalement conceptuels ou perceptifs. Selon cette théorie, les informations relatives aux événements autobiographiques sont stockées dans une hiérarchie, à différents niveaux d’abstraction (périodes de la vie, événements généraux, événements spécifiques, connaissances spécifiques aux événements ; Conway et Pleydell-Pearce, 2000 ; Conway, 2005). Il est possible que les informations épisodiques (c’est-à-dire les détails) relatives à un événement soient stockées simultanément à différents niveaux de cette structure organisationnelle, les détails conceptualisés (par exemple, » Je me souviens de cet événement avec tendresse car je tombais amoureux à ce moment précis « ) et les détails perceptifs contextualisés du même souvenir étant stockés séparément (par exemple, » Nous étions assis bras dessus bras dessous sur un banc de pique-nique à Paris » ; Conway et Pleydell-Pearce, 2000 ; Conway et al., 2016). Cette possibilité conduit à la théorie selon laquelle, en fonction de la raison pour laquelle on se souvient du passé (c’est-à-dire si les éléments conceptuels vs perceptifs sont mis en avant lors de la récupération), des processus de composantes disparates seront engagés pour activer les détails associés (Burt et al, 2003).
En d’autres termes, une personne peut adopter différentes stratégies de mémorisation pour la récupération de la mémoire autobiographique, une idée soutenue par les recherches classiques (par exemple, Schank et Abelson, 1977 ; Reiser et al., 1985) ainsi que par des résultats plus récents (Brown, 2005 ; Ford et al., 2011 ; D’Argembeau et al., 2013 ; Sheldon et Chu, 2017). Plus précisément, nous proposons qu’il existe différentes collections de processus composants qui seront activés et accentués à différents degrés pour permettre une forme conceptuelle ou perceptuelle de mémorisation. Dans la section suivante, nous proposons comment cette division cognitive se reflète également dans les modèles d’activation neuronale (Figure 1).
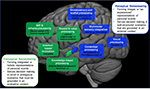
Figure 1. Une illustration des réseaux neuronaux qui soutiennent les formes conceptuelles vs perceptuelles de la remémoration autobiographique. Le réseau de mémorisation conceptuelle (représenté en vert) est proposé pour impliquer les régions du cerveau qui sont impliquées dans le traitement schématique (PFC médial dorsal et ventral), basé sur l’émotion (amygdale), basé sur la récompense et la valeur (striatum ventral), et basé sur la connaissance (cx temporal antérieur et latéral, cx perirhinal). Ce réseau est engagé par l’activation de l’hippocampe antérieur. Le réseau de mémorisation perceptive (représenté en bleu) est proposé pour impliquer les régions du cerveau qui sont impliquées dans le traitement contextuel (cx rétrosplénial, cx parahippocampique), somatosensoriel et spatial (cx somatosensoriel, précunéus antérieur), visuel (cx occipital) ainsi que les régions impliquées dans l’intégration sensorielle multimodale (lobule pariétal inférieur). Ce réseau est engagé via l’activation de l’hippocampe postérieur. Abréviations : PFC, cortex préfrontal ; cx, cortex.
Systèmes neuronaux distincts pour les formes de souvenir autobiographique épisodique
La recherche sur la mémoire autobiographique a noté des distinctions neuronales entre le souvenir de connaissances autobiographiques épisodiques (Cette fois-là, j’étais drôle) ou sémantiques (Je suis drôle) (Tulving, 1972 ; pour des exemples plus récents, voir Burianova et al, 2010 ; Brown et al., 2018), mais peu d’études se sont penchées sur les distinctions entre les différentes formes de mémoire autobiographique épisodique. La division que nous proposons entre la mémoire conceptuelle et la mémoire perceptive suppose que différents détails de mémoire épisodique d’un événement rappelé sont utilisés pour former la représentation sous-jacente, qui est reflétée dans le cerveau. Les preuves de la manière dont cette division se reflète dans le cerveau proviennent d’une division rapportée au sein du réseau par défaut – un ensemble de régions cérébrales qui se chevauchent considérablement avec le réseau de la mémoire autobiographique – qui ressemble à la mémorisation conceptuelle et perceptuelle (Buckner et al., 2008 ; Spreng et al., 2009 ; Andrews-Hanna et al., 2014). Ces recherches ont permis de décrire deux sous-systèmes corticaux du réseau par défaut qui accèdent à différents types d’informations auto-générées et les traitent. Un circuit, appelé sous-système dorsal-médian, est impliqué dans le traitement des informations conceptuelles et schématiques stockées liées aux expériences d’une personne. Le sous-système dorso-médian est composé de régions du cerveau impliquées dans le traitement évaluatif, schématique et basé sur l’essentiel, et est utilisé pour former des représentations abstraites des expériences perceptives (par exemple, Yarkoni et al., 2008 ; Binder et al., 2009 ; Binder et Desai, 2011 ; Lin et al., 2016). Un autre circuit, appelé le sous-système médio-temporal, est impliqué dans le traitement des informations perceptives et imagées générées par soi-même. Les régions cérébrales impliquées dans ce circuit sont celles impliquées dans le traitement perceptif et basé sur le contexte (ex, cortex rétrosplénial, cortex parahippocampique, lobule pariétal inférieur), ce qui permet à ce sous-système de former des représentations d’événements mentaux en réactivant ce qui a été expérimenté extérieurement (vu, entendu) pendant l’événement.
Une autre preuve neuronale d’une division neuronale entre la mémorisation conceptuelle et perceptive provient d’un modèle de mémoire qui propose des sous-systèmes similaires au réseau par défaut pour les formes de récupération de la mémoire de reconnaissance. Ce modèle propose deux sous-systèmes mnémoniques qui émergent des lobes temporaux médians (LTM) pour accéder à différents contenus épisodiques (Ranganath et Ritchey, 2012 ; Ritchey et al., 2015 ; Reagh et Ranganath, 2018). Il existe un sous-système du lobe temporal antérieur qui relie la région du MTL impliquée dans le traitement conceptuel (c’est-à-dire le cortex périrhinal) à certaines des régions trouvées dans le sous-système dorso-médian pour la récupération des connaissances conceptuelles et celles importantes pour l’évaluation des informations . Il existe également un réseau médian postérieur qui relie la région du MTL impliquée dans le traitement de l’information contextuelle externe (c.-à-d. le cortex parahippocampique) à des régions situées dans le sous-système médian-temporel qui permettent de récupérer des éléments situationnels spécifiques d’un événement rencontré (par exemple, le cortex rétrosplénial), Nous suggérons qu’une division similaire du sous-système existe pour la récupération des détails épisodiques conceptuels ou perceptuels des souvenirs autobiographiques et la recherche a déjà commencé à fournir des preuves à l’appui (Figure 1). Certaines études comparent l’activité neuronale au cours de différentes étapes de la récupération des souvenirs autobiographiques : au cours de l’accès précoce et de l’élaboration ultérieure de la mémoire autobiographique. Au cours de la phase d’accès précoce, des informations d’ordre supérieur sur un événement sont récupérées et évaluées, ce qui nécessite le « système conceptuel » de la mémoire autobiographique. Au cours de la phase d’élaboration ultérieure, les détails perceptifs et expérientiels d’un événement sont accessibles, ce qui nécessite le « système perceptif » (Addis et al., 2007 ; St. Jacques et al., 2011 ; McCormick et al., 2015). D’autres recherches sur la mémoire autobiographique se sont intéressées aux régions neuronales qui permettent de retrouver des événements autobiographiques généraux ou spécifiques, ce qui rappelle la division que nous avons proposée entre la mémoire conceptuelle et la mémoire perceptive (Addis et al., 2004 ; Levine et al., 2004). Une de ces études a rapporté que la récupération de souvenirs autobiographiques en tant qu’événements spécifiques ou en tant que connaissances personnelles activait un certain nombre de régions, y compris le MTL, mais que les événements spécifiques recrutaient des régions impliquées dans le sous-système perceptif (précuneus et lobe pariétal supérieur) ainsi que des régions d’autoréférence (PFC antéromédial ; Ford et al., 2011). Enfin, dans l’une de nos récentes expériences, nous avons directement testé comment les sous-systèmes conceptuel et perceptif soutiendraient le souvenir d’un même souvenir autobiographique de différentes manières. Nous avons mené une étude d’imagerie par résonance magnétique fonctionnelle (IRMf) dans laquelle les participants se concentraient soit sur les éléments conceptuels (les détails de la thématique ou de l’action), soit sur les éléments perceptifs (les détails visuels et contextuels de l’événement) d’un souvenir autobiographique. Notre principale conclusion était que différents réseaux neuronaux, alignés sur ce qui est présenté dans la figure 1, soutenaient de manière unique le souvenir d’un événement en tant que concept ou percept (Gurguryan et Sheldon, soumis ; pour une conclusion connexe, voir Martial et al., 2018). Dans la section suivante, nous proposons que ces réseaux à grande échelle sont systématiquement engagés par l’hippocampe antérieur et postérieur pour repérer ces détails, déterminant si un souvenir est rappelé conceptuellement ou perceptivement.
Contributions hippocampiques aux formes de remémoration autobiographique épisodique
Lors de la récupération de la mémoire autobiographique, l’hippocampe associe et intègre des informations provenant de systèmes de traitement plus larges pour accéder aux détails de la mémoire afin de former une représentation mentale cohérente (Nadel et Moscovitch, 1997 ; Hassabis et Maguire, 2009 ; Winocur et Moscovitch, 2011 ; Maguire et Mullally, 2013 ; Moscovitch et al, 2016 ; Sheldon et Levine, 2016 ; Sekeres et al., 2018). Traditionnellement, ce rôle de l’hippocampe dans la récupération a été étudié en considérant l’hippocampe comme une structure homogène, pourtant, il existe de plus en plus de preuves que l’hippocampe antérieur et postérieur remplit des fonctions distinctes de récupération de la mémoire (Poppenk et al., 2013 ; Strange et al., 2014). Dans le cas de la mémoire autobiographique, ces distinctions fonctionnelles le long de l’axe longitudinal de l’hippocampe sont souvent interprétées selon la théorie du gradient, de sorte que l’hippocampe antérieur et l’hippocampe postérieur favorisent respectivement l’accès aux détails à gros grain et aux détails à grain fin d’un souvenir (Evensmoen et al., 2013 ; Collin et al., 2015 ; McCormick et al., 2015 ; Sheldon et Levine, 2015). L’activité de l’hippocampe antérieur est censée suivre l’accès aux détails conceptuels des souvenirs personnels passés (par exemple, se souvenir qu’une conférence a eu lieu dans un hôtel en bord de mer), tandis que l’activité de l’hippocampe postérieur suit l’accès et l’élaboration des détails fins de l’événement (par exemple, se souvenir d’être assis à côté de Phife à la conférence). D’autres éléments indiquent que l’hippocampe antérieur et l’hippocampe postérieur sont réglés différemment pour représenter des informations mnémoniques nouvelles ou familières (perceptives ou expérientielles). L’hippocampe antérieur est le centre d’un réseau de nouveauté à plus grande échelle pour la mémoire et répond aux nouvelles interprétations d’anciens événements (Poppenk et al., 2010 ; Kafkas et Montaldi, 2018), tandis que l’hippocampe postérieur est situé pour répondre aux informations perceptives et expérientielles familières d’un événement (voir Kondo et al, 2008 ; McCormick et al., 2015 ; Zeidman et al., 2015).
Ces divergences dans la fonction hippocampique permettent de se souvenir d’un événement pour différentes raisons. De plus, la raison de se souvenir d’une expérience (développée dans la section suivante), signalée par les régions cérébrales préfrontales, est ce qui détermine le placement de l’activité le long de l’axe longitudinal hippocampique pour diriger la récupération de la mémoire autobiographique (Preston et Eichenbaum, 2013 ; Rajasethupathy et al., 2015). Si le but est de récupérer un souvenir autobiographique épisodique de manière conceptuelle, l’hippocampe antérieur sera activé de manière préférentielle pour recruter les détails associés via les régions connectées qui traitent les informations d’ordre supérieur ou à gros grain (par exemple, les cortex temporaux ; théorie du gradient) ainsi que les régions importantes pour le traitement évaluatif (vmPFC, striatum ventral). Ainsi, la représentation de la mémoire qui en résulte s’écarte de l’expérience d’encodage initiale (une nouvelle représentation). Si l’objectif est de retrouver un souvenir autobiographique épisodique de manière perceptive, l’hippocampe postérieur sera activé de manière à ce que les détails d’un souvenir soient réintégrés tels qu’ils ont été expérimentés initialement. Les détails perceptifs à grain plus fin (théorie du gradient) qui représentent une approximation proche de l’expérience encodée (une représentation familière) seront accessibles via des connexions directes aux régions qui traitent et intègrent les informations somatosensorielles et perceptives (par ex, parahippocampique, cortex rétrosplénial, cortex visuel et somatosensoriel).
Notre modèle suppose que l’hippocampe antérieur et postérieur – et les réseaux neuronaux plus larges – sont interconnectés, de sorte qu’il existe une interaction obligatoire entre ces systèmes de traitement lors de la construction d’une représentation de la mémoire épisodique (Sheldon et Levine, 2016 ; pour des idées connexes, voir Burke et al., 2018). En plus de suggérer qu’un souvenir autobiographique n’est pas mémorisé comme soit/ou un événement épisodique conceptuel ou perceptuel, cette idée soulève également des questions sur la directionnalité des connexions fonctionnelles entre les segments hippocampiques lors de la mémorisation. De plus en plus de travaux suggèrent que l’hippocampe antérieur joue un rôle directif dans la récupération de la mémoire par rapport à l’hippocampe postérieur, en particulier lors de la formation de la représentation d’événements mentaux complexes. Par exemple, il existe des preuves que l’hippocampe antérieur est nécessaire pour les tâches qui impliquent la construction flexible en ligne de représentations mentales, y compris les expériences autobiographiques (McCormick et al., 2015 ; Ito et Lee, 2016 ; Mack et al., 2018 ; Monge et al., 2018), mais pas les souvenirs sémantiques plus rigides qui ne nécessitent pas cette flexibilité (par exemple, Manns et al., 2003a,b ; Winocur et al., 2010). En ce qui concerne notre cadre, il se pourrait qu’une construction conceptuelle d’ordre supérieur soit un cadre nécessaire à la remémoration des représentations autobiographiques puisque la remémoration de ces événements en tant que souvenirs épisodiques nécessite toujours la manipulation de connaissances événementielles existantes (Nadel et Moscovitch, 1997 ; Moscovitch et al., 2006, 2016 ; Sekeres et al., 2018). Cette idée, cependant, est hautement spéculative et nous l’avançons pour stimuler la recherche sur la compréhension du rôle omniprésent de l’hippocampe antérieur dans les formes et les fonctions de la mémoire.
Les fonctions du souvenir autobiographique conceptuel et perceptuel
À ce stade, nous avons discuté de la façon dont notre modèle propose que les représentations épisodiques des expériences autobiographiques puissent être formées avec des informations principalement conceptuelles ou perceptuelles. Un autre élément principal de notre modèle est que la capacité de former ces différentes représentations doit servir des fonctions disparates en dehors du domaine de la mémorisation (Alea et Bluck, 2007 ; Vandermorris et al., 2013 ; Madore et al., 2016 ; Schacter et al., 2017 ; Mar et Spreng, 2018). Une fonction bien étudiée de la mémoire autobiographique est de diriger le comportement futur, ce qui inclut des tâches cognitives telles que la résolution de problèmes, la réflexion future et la prise de décision (Pillemer, 2003 ; Bar, 2009 ; Schacter, 2012).
Un exemple de cette fonction directive est la prise de décisions basées sur la mémoire : des problèmes qui nécessitent d’accéder à des informations provenant d’un souvenir passé associé. Ces problèmes de décision peuvent se présenter comme des tâches ouvertes ou fermées (Simon et al., 1987). Les tâches ouvertes sont celles dont l’issue est incertaine et/ou pour lesquelles il existe plusieurs façons d’obtenir un résultat, comme décider de la rénovation d’une maison ou de l’organisation d’une fête. Les tâches fermées sont celles dont le cheminement est défini et qui indiquent un certain résultat, par exemple un plombier qui s’appuie sur un ensemble structuré d’actions pour décider comment réparer des toilettes cassées. Une différence essentielle entre ces scénarios de décision est que les tâches ouvertes dépendent moins de la situation (c’est-à-dire de l’environnement) dans laquelle elles se déroulent que les tâches fermées. Les tâches ouvertes varient d’une situation à l’autre (la manière de rénover une maison change en fonction de la maison), tandis que les décisions fermées sont plus susceptibles de se produire de manière similaire d’une situation à l’autre (par exemple, la manière de réparer des toilettes est similaire dans toutes les salles de bains). Cette distinction est importante pour comprendre quand la mémorisation perceptive par rapport à la mémorisation conceptuelle sera la plus efficace pour guider la prise de décision.
Puisque les décisions fermées sont plus structurées et liées à l’environnement, la mémorisation perceptive sera efficace pour utiliser les informations de l’environnement actuel d’une personne comme un indice pour accéder à un souvenir passé pertinent. Ceci est utile pour prendre des décisions rapides concernant un stimulus rencontré (par exemple, « Cet animal ou cet aliment est-il sûr ou dangereux ? »), des tâches de navigation (se rappeler le chemin précis pour aller d’un point A à un point B) ou se rappeler l’emplacement d’un objet perdu (par exemple, « Où ai-je mis mes clés ? »). L’utilisation du souvenir perceptif pour ces décisions recrutera l’hippocampe postérieur pour activer les processus perceptifs et basés sur l’expérience afin de réintégrer mentalement une expérience passée et de l’appliquer à la situation actuelle. Le souvenir perceptuel, cependant, sera moins utile pour les décisions ouvertes. En effet, les indices externes (perceptuels) provenant de l’environnement actuel d’une personne peuvent ne pas réactiver le bon souvenir passé pour rassembler les informations nécessaires à la prise de décision. Dans ces cas, la mémorisation conceptuelle est mieux adaptée car elle permet à une personne d’accéder à des représentations de mémoire généralisées et de les évaluer dans la mesure où elles s’appliquent à la nouvelle situation de décision ouverte. Des exemples de telles décisions sont des problèmes nouveaux et » bruyants » qui nécessitent d’abord de générer un résultat souhaité (par exemple, » Comment devrais-je redécorer cette salle de bain ? « ), puis d’utiliser ce but/résultat représenté en interne (concept) pour retrouver des souvenirs passés pertinents (par exemple, un autre projet de rénovation de maison entrepris dans le passé). Dans certains de nos travaux précédents, nous avons constaté que les problèmes sociaux ouverts (par exemple, se faire de nouveaux amis) nécessitent une simulation épisodique pour construire des solutions à ces problèmes (Sheldon et al., 2011 ; Vandermorris et al., 2013), ce que nous supposons être basé sur la mémorisation conceptuelle créant de nouveaux résultats à ces problèmes. L’utilisation de la remémoration conceptuelle fera appel à l’hippocampe antérieur pendant la remémoration, ce qui stimulera l’activité des régions du cerveau impliquées dans les processus schématiques et évaluatifs (par exemple, vmPFC ; Euston et al, 2012).
Bien que certaines situations puissent bénéficier de la représentation de notre passé sous forme de concepts ou de percepts principalement pour les fonctions directives de la mémoire autobiographique, la capacité à passer dynamiquement d’une forme de souvenir à une autre est probablement ce qui sous-tend l’utilisation optimale de la mémoire (voir également, Richards et Frankland, 2017 ; Duncan et Schlichting, 2018). Cette idée devient claire lorsqu’on pense aux erreurs potentielles lors de la prise de décision qui surviendraient si une seule méthode de mémorisation était utilisée, ce qui est illustré dans la figure 2. Si l’on suit cette figure, le fait de s’appuyer principalement sur la mémorisation conceptuelle peut conduire à appliquer les souvenirs autobiographiques de manière trop large (c’est-à-dire libérale), car d’autres détails pertinents d’une expérience sont ignorés (par exemple, la rencontre d’un individu aux cheveux courts et à la barbe lors d’une conférence qui racontait des blagues amusantes). Cela peut conduire à l’application incorrecte d’informations tirées d’expériences passées à un scénario de prise de décision actuel et à un résultat médiocre (par exemple, identifier par erreur d’autres personnes drôles, aux cheveux courts et barbus comme la personne de la conférence). D’autre part, une confiance excessive dans la mémorisation perceptive peut entraîner une application trop conservatrice (c’est-à-dire rigide) des connaissances autobiographiques, ce qui conduit également à des erreurs de prise de décision. Si une personne recherche des informations sur une expérience passée pour prendre une décision en se concentrant uniquement sur des détails perceptifs spécifiques, elle risque de ne pas être capable de localiser correctement un souvenir pour éclairer sa décision (par exemple, décider que la personne drôle à laquelle vous parlez, que vous avez rencontrée lors d’une conférence précédente, n’est pas cette personne parce qu’elle a changé de coiffure). En d’autres termes, l’adhésion rigide à une seule forme de mémorisation peut conduire à l’application incorrecte d’informations du passé à un scénario de prise de décision actuel. Au contraire, c’est en modifiant les contributions relatives de la façon dont nous nous souvenons que les souvenirs autobiographiques peuvent remplir plusieurs fonctions adaptatives. Compte tenu du rôle central de l’hippocampe dans ces formes de mémorisation épisodique, il est probable que cette région du cerveau soit la clé de cette adaptabilité.

Figure 2. Schéma décrivant un exemple de scénarios de prise de décision dans lesquels l’utilisation des seules formes conceptuelles ou perceptives de mémorisation conduit à un résultat correct (mais incertain) ou défectueux. Dans cet exemple, un « décideur » a rencontré un drôle d’individu aux cheveux courts et à la barbe, nommé Alan, lors d’une conférence (panneau de gauche). Dans un scénario, le décideur rencontre plus tard quelqu’un qui ressemble à Alan mais avec une coiffure différente (rangée du haut). Le décideur doit utiliser le souvenir qu’il a d’Alan lors de l’événement autobiographique pour décider si l’individu rencontré est lui. S’il se souvient de l’événement autobiographique par le biais de la mémoire perceptive, en retrouvant de manière rigide chaque détail de l’apparence d’Alan, il ne pourra pas identifier correctement cet individu comme étant Alan car sa coiffure (c’est-à-dire un détail perceptif) a changé. S’ils se souviennent de l’événement autobiographique par le biais de la mémoire conceptuelle, ils auront plus de chances de l’identifier comme étant Alan, même avec sa nouvelle coiffure ; cependant, cette décision manquera de certitude (c’est-à-dire que c’est peut-être Alan). Dans un autre scénario, le décideur rencontre plus tard un homme portant une barbe et une coiffure similaires à celles d’Alan (rangée du bas). Comme ci-dessus, s’il se souvient de sa rencontre avec Alan par le biais de la mémorisation perceptive, il décidera correctement que ce nouvel homme n’est pas Alan. S’ils se souviennent de cet événement de manière conceptuelle (c’est-à-dire en se rappelant seulement qu’ils ont rencontré un homme drôle la conférence), ils peuvent accidentellement identifier ce nouvel homme comme Alan en raison d’une caractéristique de chevauchement (être drôle) représentée au niveau du concept.
Dans cette dernière section, nous avons souligné comment notre cadre décrivant différentes formes de mémorisation impactent la prise de décision basée sur la mémoire, cependant nous prédisons que cet impact se présenterait de manière similaire pour d’autres fonctions directives de la mémoire autobiographique, y compris la planification de comportements futurs et la résolution de problèmes personnels. En dehors des fonctions directives de la mémoire autobiographique, il y a celles qui se rapportent au soi et aux fonctions sociales. Les recherches existantes ont fourni des points de vue sur la façon dont l’accès aux souvenirs autobiographiques à différents niveaux, similaires à la mémoire conceptuelle et perceptuelle, peut bénéficier et nuire à ces fonctions (par exemple, Pillemer, 2003 ; Alea et Bluck, 2007 ; Prebble et al., 2013). Bien que cela dépasse le cadre de cet article, il est intéressant de poursuivre les recherches visant à comprendre comment différentes formes de mémorisation opèrent à travers ces fonctions.
Conclusions
Les souvenirs autobiographiques sont des constructions complexes qui englobent un riche éventail d’informations, y compris des détails épisodiques conceptuels et perceptifs. Une même expérience passée peut être représentée lors de la récupération de diverses manières, selon la façon dont on accède à ces détails, et cela est déterminé par l’objectif de la remémoration. Nous avons présenté ici un bref aperçu des comptes rendus théoriques et des résultats empiriques sur l’organisation et la récupération de la mémoire autobiographique afin de suggérer une nouvelle division dans la mémoire autobiographique épisodique. Nous avons proposé deux formes de souvenir du passé – en tant que concept ou en tant que percept – et fourni un compte-rendu neuronal pour ces différentes formes de souvenir, émergeant de réseaux hippocampiques-corticaux disparates. Nous avons défini la raison d’être de ces formes de souvenir en décrivant leurs rôles fonctionnels dans la prise de décision, offrant une nouvelle perspective sur la façon dont les objectifs d’une tâche actuelle bénéficient de la flexibilité de la mémoire autobiographique épisodique. Enfin, nous avons proposé que la capacité de passer d’une forme de souvenir à une autre, spécifiée par la contribution relative des réseaux hippocampiques-corticaux décrits, est la clé de la mémoire adaptative.
Contributions de l’auteur
SS a construit la perspective présentée et le plan de l’article. SS, LG et CF ont contribué à la rédaction de la version finale et ont fourni des commentaires rédactionnels.
Funding
Le financement a été assuré par une subvention à la découverte du Conseil de recherches en sciences naturelles et en génie du Canada (#RGPIN-04241) et par des fonds du Programme des chaires de recherche du Canada, tous deux attribués à SS.
Déclaration de conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de toute relation commerciale ou financière qui pourrait être interprétée comme un conflit d’intérêts potentiel.
Remerciements
Nous tenons à remercier les présentateurs (Brian Levine, Paul Frankland et Melanie Sekeres) des membres du symposium (L’interaction dynamique entre la mémorisation et l’oubli détaillés) de la Conférence internationale sur l’apprentissage et la mémoire 2018, car la pièce a été inspirée par des conversations tenues lors de cette session.
Addis, D. R., McIntosh, A. R., Moscovitch, M., Crawley, A. P., et McAndrews, M. P. (2004). Characterizing spatial and temporal features of autobiographical memory retrieval networks : a partial least squares approach. Neuroimage 23, 1460-1471. doi : 10.1016/j.neuroimage.2004.08.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Addis, D. R., Wong, A. T., et Schacter, D. L. (2007). Se souvenir du passé et imaginer le futur : substrats neuronaux communs et distincts pendant la construction et l’élaboration des événements. Neuropsychologia 45, 1363-1377. doi : 10.1016/j.neuropsychologia.2006.10.016
PubMed Abstract | CrossRef Full Text | Google Scholar
Alea, N., et Bluck, S. (2007). Je te garderai à l’esprit : la fonction d’intimité de la mémoire autobiographique. Appl. Cogn. Psychol. 21, 1091-1111. doi : 10.1002/acp.1316
CrossRef Full Text | Google Scholar
Andrews-Hanna, J. R., Smallwood, J., and Spreng, R. N. (2014). Le réseau par défaut et la pensée auto-générée : processus de composants, contrôle dynamique et pertinence clinique. Ann. N. Y. Acad. Sci. 1316, 29-52. doi : 10.1111/nyas.12360
PubMed Abstract | CrossRef Full Text | Google Scholar
Bar, M. (2009). Le cerveau proactif : la mémoire des prédictions. Philos. Trans. R. Soc. Lond. B Biol. Sci. 364, 1235-1243. doi : 10.1098/rstb.2008.0310
PubMed Abstract | CrossRef Full Text | Google Scholar
Bartlett, F. C. (1932). Se souvenir : une étude expérimentale et sociale. Cambridge : Cambridge University.
Google Scholar
Binder, J. R., et Desai, R. H. (2011). La neurobiologie de la mémoire sémantique. Trends Cogn. Sci. 15, 527-536. doi : 10.1016/j.tics.2011.10.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Binder, J. R., Desai, R. H., Graves, W. W., et Conant, L. L. (2009). Où est le système sémantique ? Une revue critique et une méta-analyse de 120 études de neuroimagerie fonctionnelle. Cereb. Cortex 19, 2767-2796. doi : 10.1093/cercor/bhp055
PubMed Abstract | CrossRef Full Text | Google Scholar
Brown, N. R. (2005). Sur la prévalence des groupes d’événements dans la mémoire autobiographique. Soc. Cogn. 23, 35-69. doi : 10.1521/soco.23.1.35.59194
CrossRef Full Text | Google Scholar
Brown, T. I., Rissman, J., Chow, T. E., Uncapher, M. R., and Wagner, A. D. (2018). Contributions différentielles du lobe temporal médian et du cortex pariétal à la mémoire épisodique autobiographique du monde réel et à la mémoire sémantique autobiographique. Sci. Rep. 8:6190. doi : 10.1038/s41598-018-24549-y
PubMed Abstract | CrossRef Full Text | Google Scholar
Buckner, R. L., Andrews-Hanna, J. R., et Schacter, D. L. (2008). Le réseau par défaut du cerveau : anatomie, fonction et pertinence pour la maladie. Ann. N. Y. Acad. Sci. 1124, 1-38. doi : 10.1196/annals.1440.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Burianova, H., McIntosh, A. R., et Grady, C. L. (2010). Un réseau cérébral fonctionnel commun pour la récupération de la mémoire autobiographique, épisodique et sémantique. Neuroimage 49, 865-874. doi : 10.1016/j.neuroimage.2009.08.066
PubMed Abstract | CrossRef Full Text | Google Scholar
Burke, S. N., Gaynor, L. S., Barnes, C. A., Bauer, R. M., Bizon, J. L., Roberson, E. D., et al. (2018). Fonctions partagées des cortex périrhinal et parahippocampique : implications pour le vieillissement cognitif. Trends Neurosci. 41, 349-359. doi : 10.1016/j.tins.2018.03.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Burt, C. D. B., Kemp, S., et Conway, M. A. (2003). Thèmes, événements et épisodes dans la mémoire autobiographique. Mem. Cognit. 31, 317-325. doi : 10.3758/bf03194390
PubMed Abstract | CrossRef Full Text | Google Scholar
Cabeza, R., et Moscovitch, M. (2013). Systèmes de mémoire, modes de traitement et composants : preuves de neuroimagerie fonctionnelle. Perspect. Psychol. Sci. 8, 49-55. doi : 10.1177/1745691612469033
PubMed Abstract | CrossRef Full Text | Google Scholar
Collin, S. H. P., Milivojevic, B., et Doeller, C. F. (2015). Les hiérarchies de mémoire se cartographient sur le grand axe hippocampique chez l’homme. Nat. Neurosci. 18, 1562-1564. doi : 10.1038/nn.4138
PubMed Abstract | CrossRef Full Text | Google Scholar
Conway, M. A. (2001). La mémoire épisodique sensorielle-perceptuelle et son contexte : la mémoire autobiographique. Philos Trans. R. Soc. Lond. B Biol. Sci. 356, 1375-1384. doi : 10.1098/rstb.2001.0940
PubMed Abstract | CrossRef Full Text | Google Scholar
Conway, M. A. (2005). Memory and the self. J. Mem. Lang. 53, 594-628. doi : 10.1016/j.jml.2005.08.005
CrossRef Full Text | Google Scholar
Conway, M. A., Loveday, C., et Cole, S. N. (2016). Le système de mémorisation-imagination. Mem. Stud. 9, 256-265. doi : 10.1177/1750698016645231
CrossRef Full Text | Google Scholar
Conway, M. A., et Pleydell-Pearce, C. W. (2000). La construction des souvenirs autobiographiques dans le système de mémoire de soi. Psychol. Rev. 107, 261-288. doi : 10.1037//0033-295x.107.2.261
PubMed Abstract | CrossRef Full Text | Google Scholar
D’Argembeau, A., Cassol, H., Phillips, C., Balteau, E., Salmon, E., et Van der Linden, M. (2013). Les cerveaux créant des histoires de soi : la base neuronale du raisonnement autobiographique. Soc. Cogn. Affect. Neurosci. 9, 646-652. doi : 10.1093/scan/nst028
PubMed Abstract | CrossRef Full Text | Google Scholar
Duncan, K. D., et Schlichting, M. L. (2018). Représentations hippocampiques en fonction du temps, de la sous-région et de l’état du cerveau. Neurobiol. Learn. Mem. 153, 40-56. doi : 10.1016/j.nlm.2018.03.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Euston, D. R., Gruber, A. J., et McNaughton, B. L. (2012). Le rôle du cortex préfrontal médian dans la mémoire et la prise de décision. Neuron 76, 1057-1070. doi : 10.1016/j.neuron.2012.12.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Evensmoen, H. R., Lehn, H., Xu, J., Witter, M. P., Nadel, L., et Håberg, A. K. (2013). L’hippocampe antérieur soutient une représentation globale et grossière de l’environnement et l’hippocampe postérieur soutient des représentations locales et à grain fin de l’environnement. J. Cogn. Neurosci. 11, 1908-1925. doi : 10.1162/jocn_a_00436
PubMed Abstract | CrossRef Full Text | Google Scholar
Ford, J. H., Addis, D. R., et Giovanello, K. S. (2011). Activité neuronale différentielle pendant la recherche de souvenirs autobiographiques spécifiques et généraux suscités par des indices musicaux. Neuropsychologia 49, 2514-2526. doi : 10.1016/j.neuropsychologia.2011.04.032
PubMed Abstract | CrossRef Full Text | Google Scholar
Greenberg, D. L., et Rubin, D. C. (2003). La neuropsychologie de la mémoire autobiographique. Cortex 39, 687-728. doi : 10.1016/s0010-9452(08)70860-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Hassabis, D., et Maguire, E. A. (2009). Le système de construction du cerveau. Philos. Trans. R. Soc. Lond. B Biol. Sci. 364, 1263-1271. doi : 10.1098/rstb.2008.0296
PubMed Abstract | CrossRef Full Text | Google Scholar
Ito, R., et Lee, A. C. H. (2016). Le rôle de l’hippocampe dans la prise de décision de conflit d’approche-évitement : preuves des études sur les rongeurs et les humains. Behav. Brain Res. 313, 345-357. doi : 10.1016/j.bbr.2016.07.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Kafkas, A., et Montaldi, D. (2018). L’attente affecte l’apprentissage et module l’expérience de la mémoire lors de la récupération. Cognition 180, 123-134. doi : 10.1016/j.cognition.2018.07.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Kondo, H., Lavenex, P., et Amaral, D. G. (2008). Connexions intrinsèques de la formation hippocampique du singe macaque : I. Le gyrus denté. J. Comp. Neurol. 511, 497-520. doi : 10.1002/cne.21825
PubMed Abstract | CrossRef Full Text | Google Scholar
Levine, B., Turner, G. R., Tisserand, D., Hevenor, S. J., Graham, S. J., and McIntosh, A. R. (2004). The functional neuroanatomy of episodic and semantic autobiographical remembering : a prospective functional MRI study. J. Cogn. Neurosci. 16, 1633-1646. doi : 10.1162/0898929042568587
PubMed Abstract | CrossRef Full Text | Google Scholar
Lin, W.-J., Horner, A. J., et Burgess, N. (2016). Le cortex préfrontal ventromédial, une valeur ajoutée aux souvenirs autobiographiques. Sci. Rep. 6:28630. doi : 10.1038/srep28630
PubMed Abstract | CrossRef Full Text | Google Scholar
Mack, M. L., Love, B. C., et Preston, A. R. (2018). Construire des concepts un épisode à la fois : l’hippocampe et la formation de concepts. Neurosci. Lett. 680, 31-38. doi : 10.1016/j.neulet.2017.07.061
PubMed Abstract | CrossRef Full Text | Google Scholar
Madore, K. P., Jing, H. G., et Schacter, D. L. (2016). Pensée créative divergente chez les jeunes et les adultes plus âgés : extension des effets d’une induction de spécificité épisodique. Mem. Cognit. 44, 974-988. doi : 10.3758/s13421-016-0605-z
PubMed Abstract | CrossRef Full Text | Google Scholar
Maguire, E. A., et Mullally, S. L. (2013). L’hippocampe : un manifeste pour le changement. J. Exp. Psychol. Gen. 142, 1180-1189. doi : 10.1037/a0033650
PubMed Abstract | CrossRef Full Text | Google Scholar
Manns, J. R., Hopkins, R. O., Reed, J. M., Kitchener, E. G., and Squire, L. R. (2003a). Recognition memory and the human hippocampus. Neuron 37, 171-180. doi : 10.1016/S0896-6273(02)01147-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Manns, J. R., Hopkins, R. O., and Squire, L. R. (2003b). La mémoire sémantique et l’hippocampe humain. Neuron 38, 127-133. doi : 10.1016/s0896-6273(03)00146-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Mar, R. A., et Spreng, R. N. (2018). La mémoire épisodique résout à la fois des problèmes sociaux et non sociaux et a évolué pour remplir de nombreuses fonctions différentes. Behav. Brain Sci. 41:e20. doi : 10.1017/s0140525x17001418
PubMed Abstract | CrossRef Full Text | Google Scholar
Martial, C., Stawarczyk, D., et D’Argembeau, A. (2018). Corrélats neuronaux de la connaissance de soi indépendante du contexte et dépendante du contexte. Cerveau et cognition. 125, 23-31. doi : 10.1016/j.bandc.2018.05.004
PubMed Abstract | CrossRef Full Text | Google Scholar
McCormick, C., St-Laurent, M., Ty, A., Valiante, T. A., et McAndrews, M. P. (2015). Connectivité fonctionnelle et effective hippocampique-néocorticale pendant la construction et l’élaboration de la récupération de la mémoire autobiographique. Cereb. Cortex 25, 1297-1305. doi : 10.1093/cercor/bht324
PubMed Abstract | CrossRef Full Text | Google Scholar
Monge, Z. A., Wing, E. A., Stokes, J., et Cabeza, R. (2018). Recherche et récupération de souvenirs autobiographiques et de laboratoire : composants neuronaux partagés et distincts. Neuropsychologia 110, 44-54. doi : 10.1016/j.neuropsychologia.2017.07.030
PubMed Abstract | CrossRef Full Text | Google Scholar
Moscovitch, M., Cabeza, R., Winocur, G., et Nadel, L. (2016). La mémoire épisodique et au-delà : l’hippocampe et le néocortex en transformation.Annu. Rev. Psychol. 67, 105-134. doi : 10.1146/annurev-psych-113011-143733
PubMed Abstract | CrossRef Full Text | Google Scholar
Moscovitch, M., Nadel, L., Winocur, G., Gilboa, A., et Rosenbaum, R. S. (2006). La neuroscience cognitive de la mémoire épisodique, sémantique et spatiale à distance. Curr. Opin. Neurobiol. 16, 179-190. doi : 10.1016/j.conb.2006.03.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Nadel, L., et Moscovitch, M. (1997). Consolidation de la mémoire, amnésie rétrograde et complexe hippocampique. Curr. Opin. Neurobiol. 7, 217-227. doi : 10.1016/s0959-4388(97)80010-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Pillemer, D. B. (2003). Fonctions directives de la mémoire autobiographique : le pouvoir de guidage de l’épisode spécifique. Memory 11, 193-202. doi : 10.1080/741938208
PubMed Abstract | CrossRef Full Text | Google Scholar
Poppenk, J., Evensmoen, H. R., Moscovitch, M., et Nadel, L. (2013). Spécialisation de l’axe long de l’hippocampe humain. Trends Cogn. Sci. 17, 230-240. doi : 10.1016/j.tics.2013.03.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Poppenk, J., Köhler, S., et Moscovitch, M. (2010). Revisiter l’effet de nouveauté : Quand la familiarité, et non la nouveauté, améliore la mémoire. J. Exp. Psychol. Learn. Mem. Cogn. 36, 1321-1330. doi : 10.1037/a0019900
PubMed Abstract | CrossRef Full Text | Google Scholar
Prebble, S. C., Addis, D. R., et Tippett, L. J. (2013). La mémoire autobiographique et le sens du soi. Psychol. Bull. 139, 815-840. doi : 10.1037/a0030146
PubMed Abstract | CrossRef Full Text | Google Scholar
Preston, A. R., et Eichenbaum, H. (2013). Interaction de l’hippocampe et du cortex préfrontal dans la mémoire. Curr. Biol. 23, R764-R773. doi : 10.1016/j.cub.2013.05.041
PubMed Abstract | CrossRef Full Text | Google Scholar
Rajasethupathy, P., Sankaran, S., Marshel, J. H., Kim, C. K., Ferenczi, E., Lee, S. Y., et al. (2015). Les projections du néocortex médient le contrôle descendant de la récupération de la mémoire. Nature 526, 653-659. doi : 10.1038/nature15389
PubMed Abstract | CrossRef Full Text | Google Scholar
Ranganath, C., et Ritchey, M. (2012). Deux systèmes corticaux pour le comportement guidé par la mémoire. Nat. Rev. Neurosci. 13, 713-726. doi : 10.1038/nrn3338
PubMed Abstract | CrossRef Full Text | Google Scholar
Reagh, Z. M., et Ranganath, C. (2018). Que nous dit l’organisation fonctionnelle des réseaux cortico-hippocampiques sur l’organisation fonctionnelle de la mémoire ? Neurosci. Lett. 680, 69-76. doi : 10.1016/j.neulet.2018.04.050
PubMed Abstract | CrossRef Full Text | Google Scholar
Reiser, B. J., Black, J. B., et Abelson, R. P. (1985). Structures de connaissances dans l’organisation et la récupération des souvenirs autobiographiques. Cogn. Psychol. 17, 89-137. doi : 10.1016/0010-0285(85)90005-2
CrossRef Full Text | Google Scholar
Richards, B. A., and Frankland, P. W. (2017). La persistance et la fugacité de la mémoire. Neuron 94, 1071-1084. doi : 10.1016/j.neuron.2017.04.037
PubMed Abstract | CrossRef Full Text | Google Scholar
Ritchey, M., Libby, L. A., Ranganath, C. (2015). Systèmes cortico-hippocampiques impliqués dans la mémoire et la cognition : le cadre PMAT. Prog. Brain Res. 219, 45-64. doi : 10.1016/bs.pbr.2015.04.001
CrossRef Full Text | Google Scholar
Rubin, D. C. (2005). Une approche des systèmes de base à la mémoire autobiographique. Curr. Dir. Psychol. Sci. 14, 79-83. doi : 10.1111/j.0963-7214.2005.00339.x
CrossRef Full Text | Google Scholar
Rubin, D. C. (2006). Le modèle des systèmes de base de la mémoire épisodique. Perspect. Psychol. Sci. 1, 277-311. doi : 10.1111/j.1745-6916.2006.00017.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Schacter, D., et Addis, D. R. (2007). La neuroscience cognitive de la mémoire constructive : se souvenir du passé et imaginer l’avenir. Philos. Trans. R. Soc. Lond. B Biol. Sci. 362, 773-786. doi : 10.1098/rstb.2007.2087
PubMed Abstract | CrossRef Full Text | Google Scholar
Schacter, D. L. (2012). Les processus constructifs adaptatifs et l’avenir de la mémoire. Am. Psychol. 67, 603-613. doi : 10.1037/a0029869
PubMed Abstract | CrossRef Full Text | Google Scholar
Schacter, D. L., Benoit, R. G., et Szpunar, K. K. (2017). La pensée future épisodique : mécanismes et fonctions. Curr. Opin. Behav. Sci. 17, 41-50. doi : 10.1016/j.cobeha.2017.06.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Schacter, D. L., Guerin, S. A., et St. Jacques, P. L. (2011). La distorsion de la mémoire : une perspective adaptative. Trends Cogn. Sci. 15, 467-474. doi : 10.1016/j.tics.2011.08.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Schank, R. C., et Abelson, R. P. (1977). Scénarios, plans, objectifs et compréhension. New York, NY : Psychology Press.
Google Scholar
Sekeres, M. J., Winocur, G., et Moscovitch, M. (2018). L’hippocampe et les structures néocorticales connexes dans la transformation de la mémoire. Neurosci. Lett. 680, 39-53. doi : 10.1016/j.neulet.2018.05.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Sheldon, S., et Chu, S. (2017). Quoi versus où : enquête sur la façon dont la récupération de la mémoire autobiographique diffère lorsqu’on y accède avec des informations thématiques versus spatiales. Q. J. Exp. Psychol. 70, 1909-1921. doi : 10.1080/17470218.2016.1215478
PubMed Abstract | CrossRef Full Text | Google Scholar
Sheldon, S., et Levine, B. (2015). Les lobes temporaux médians distinguent les relations intra-item et item-contexte pendant la récupération de la mémoire autobiographique. Hippocampe 1590, 1577-1590. doi : 10.1002/hipo.22477
PubMed Abstract | CrossRef Full Text | Google Scholar
Sheldon, S., et Levine, B. (2016). Le rôle de l’hippocampe dans la mémoire et la construction mentale. Ann. N. Y. Acad. Sci. 1369, 76-92. doi : 10.1111/nyas.13006
PubMed Abstract | CrossRef Full Text | Google Scholar
Sheldon, S., McAndrews, M. P., et Moscovitch, M. (2011). Les processus de mémoire épisodique médiés par les lobes temporaux médians contribuent à la résolution de problèmes ouverts. Neuropsychologia 49, 2439-2447. doi : 10.1016/j.neuropsychologia.2011.04.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Simon, H. A., Dantzig, G. B., Hogarth, R., Plott, C. R., Raiffa, H., Schelling, T. C., et al. (1987). Decision making and problem solving. Interfaces 17, 11-31. doi : 10.1287/inte.17.5.11
CrossRef Full Text | Google Scholar
Spreng, R. N., Mar, R. A., et Kim, A. S. N. (2009). La base neuronale commune de la mémoire autobiographique, de la prospection, de la navigation, de la théorie de l’esprit et du mode par défaut : une méta-analyse quantitative. J. Cogn. Neurosci. 21, 489-510. doi : 10.1162/jocn.2008.21029
PubMed Abstract | CrossRef Full Text | Google Scholar
St. Jacques, P. L., Kragel, P. A., and Rubin, D. C. (2011). Réseaux neuronaux dynamiques soutenant la récupération de la mémoire. Neuroimage 57, 608-616. doi : 10.1016/j.neuroimage.2011.04.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Strange, B. A., Witter, M. P., Lein, E. S., et Moser, E. I. (2014). Organisation fonctionnelle de l’axe longitudinal de l’hippocampe. Nat. Rev. Neurosci. 15, 655-669. doi : 10.1038/nrn3785
PubMed Abstract | CrossRef Full Text | Google Scholar
Szpunar, K. K., et McDermott, K. B. (2009). « Episodic future thought : remembering the past to imagine the future, » in Handbook of Imagination and Mental Simulation, eds K. D. Markman, W. M. P. Klein, J. A. Suhr (New York, NY : Psychology Press), 119-129.
Google Scholar
Tulving, E. (1972). La mémoire épisodique et la mémoire sémantique. Organ. Memory 1, 381-403.
Google Scholar
Tulving, E. (2002). La mémoire épisodique : de l’esprit au cerveau. Annu. Rev. Psychol. 53, 1-25. doi : 10.1146/annurev.psych.53.100901.135114
PubMed Abstract | CrossRef Full Text | Google Scholar
Vandermorris, S., Sheldon, S., Winocur, G., et Moscovitch, M. (2013). Contributions différentielles des fonctions de mémoire exécutive et épisodique à la résolution de problèmes chez les adultes plus jeunes et plus âgés. J. Int. Neuropsychol. Soc. 19, 1087-1096. doi : 10.1017/s1355617713000982
PubMed Abstract | CrossRef Full Text | Google Scholar
Winocur, G., et Moscovitch, M. (2011). Transformation de la mémoire et consolidation des systèmes. J. Int. Neuropsychol. Soc. 17, 766-780. doi : 10.1017/S1355617711000683
PubMed Abstract | CrossRef Full Text | Google Scholar
Winocur, G., Moscovitch, M., et Bontempi, B. (2010). Formation de la mémoire et rétention à long terme chez l’homme et l’animal : Convergence vers un compte de transformation des interactions hippocampique-néocortical. Neuropsychologia 48, 2339-2356. doi : 10.1016/j.neuropsychologia.2010.04.016
PubMed Abstract | CrossRef Full Text | Google Scholar
Yarkoni, T., Speer, N. K., et Zacks, J. M. (2008). Substrats neuronaux de la compréhension et de la mémoire narrative. Neuroimage 41, 1408-1425. doi : 10.1016/j.neuroimage.2008.03.062
PubMed Abstract | CrossRef Full Text | Google Scholar
Zeidman, P., Mullally, S. L., et Maguire, E. A. (2015). Construire, percevoir et maintenir des scènes : activité et connectivité hippocampiques. Cereb. Cortex 25, 3836-3855. doi : 10.1093/cercor/bhu266
PubMed Abstract | CrossRef Full Text | Google Scholar
.