
Isolation et caractérisation des dérivés de la ptérosine de P. aquilinum
Des dérivés de la ptérosine ont été isolés à partir des plantes entières de P. aquilinum. Pour l’étude des constituants phytochimiques de la fougère fougère, un extrait aqueux a été successivement partitionné avec de l’acétate d’éthyle (EtOAc) et du n-butanol (n-BuOH) (Fig. 1). La chromatographie sur colonne répétée de la fraction soluble dans l’EtOAc a permis d’isoler neuf dérivés, dont la (2R)-ptérosine B (1), la ptérosine Z (2), la (2S)-ptérosine P (3), (3R)-ptérosine D (4), (2S)-ptérosine A (5), (2S,3R)-ptérosine C (6), (2R,3R)-ptérosine C (7), (2R)-ptéroside B (8), et ptéroside Z (9), avec des puretés supérieures à 97% (Fig. 2). La séparation chromatographique répétée sur colonne de la fraction soluble dans le n-BuOH a permis d’isoler six dérivés, dont le (2S)-ptéroside A (10), le (2S)-ptéroside A2 (11), le (2S,3R)-ptéroside C (12), le (2R,3R)-ptéroside C (13), le (3S)-ptéroside D (14) et le (2S)-ptéroside P (15), avec des puretés supérieures à 97 %. Les structures des composés ont été identifiées par l’analyse des données spectrales, notamment MS, 1D- et 2D-NMR (Informations supplémentaires 1).

Structures des composés de ptérosine 1-15xxx
Activité inhibitrice des dérivés de ptérosine contre BACE1, AChE, et BChE
Pour évaluer le potentiel anti-AD, l’activité inhibitrice de chaque composé de ptérosine contre BACE1 et les cholinestérases a été évaluée par des essais d’inhibition in vitro respectifs (Tableau 1). Tous les dérivés de la ptérosine testés ont montré des activités inhibitrices concentration-dépendantes contre BACE1 avec une gamme de valeurs IC50 (concentration inhibitrice demi-maximale) de 9,74-94,4 μM, à l’exception de la (2S)-ptérosine A et du (2S)-ptéroside P qui étaient inactifs aux concentrations testées, par rapport à l’IC50 de la quercétine utilisée comme contrôle positif, qui était de 18,8 μM. Le pouvoir inhibiteur des inhibiteurs les plus puissants était dans l’ordre le (2R,3R)-ptéroside C, le (3S)-ptéroside D, le (2R)-ptéroside B, la (2S,3R)-ptérosine C, la (2R,3R)-ptérosine C, le (2S,3R)-ptéroside C et la (2R)-ptérosine B avec des valeurs de CI50 de 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 et 29,6 μM, respectivement. Nous avons ensuite testé les potentiels inhibiteurs des dérivés de la ptérosine contre l’AChE. Tous les composés testés ont montré des activités inhibitrices significatives de l’AChE, avec des valeurs IC50 comprises entre 2,55 et 110 μM, par rapport à l’IC50 contre l’AChE de la berbérine utilisée comme contrôle positif, qui était de 0,39 μM. Les composés de ptérosine qui ont présenté la plus forte activité inhibitrice contre l’AChE étaient le (2R)-ptéroside B, le (2R,3R)-ptéroside C, le (2S,3R)-ptéroside C, le (2S,3R)-ptérosine C et le (2R)-ptérosine B avec des valeurs de CI50 de 2,55, 3,77, 9,17, 12,8 et 16,2 μM, respectivement. Enfin, nous avons testé les capacités inhibitrices des dérivés de la ptérosine contre le BChE. Tous les composés de ptérosine testés ont montré une activité inhibitrice contre BChE, avec des valeurs de CI50 qui variaient de 5,29 à 119 μM, à l’exception de la (3R)-ptérosine D qui était inactive aux concentrations testées, comparée à la CI50 de la berbérine contre BChE, qui était de 3,32 μM. Les composés de ptérosine qui ont présenté la plus forte activité inhibitrice contre BChE étaient le (2R,3R)-ptéroside C et le ptéroside Z avec des valeurs de CI50 de 5,29 et 5,31 μM, respectivement.
Collectivement, la plupart des dérivés de la ptérosine testés ont présenté des activités inhibitrices significatives contre BACE1, AChE, et BChE simultanément. La présence du groupe supplémentaire 2-hydroxyméthyl-tétrahydro-pyran-3,4,5-triol comme dans les dérivés de la ptérosine a augmenté de manière significative les activités inhibitrices contre les enzymes. De plus, la présence du groupe hydroxyméthyle supplémentaire en position 2 du cycle indanone de la (2R)-ptérosine B comme dans la (2S)-ptérosine A ou du groupe méthyle comme dans la (3R)-ptérosine D et la ptérosine Z a diminué les activités inhibitrices contre les enzymes. En particulier, la présence du groupe hydroxyméthyle en position-5 du cycle indanone comme dans la (2S)-ptérosine P a diminué l’activité inhibitrice contre la BACE1.
Paramètres cinétiques de l’inhibition enzymatique par les dérivés de la ptérosine
Pour tenter d’expliquer le mode d’inhibition enzymatique des dérivés de la ptérosine, nous avons réalisé une analyse cinétique de la BACE1 et des cholinestérases pour des inhibiteurs représentatifs (tableau 2, informations complémentaires 2). Un Ki (constante d’inhibition) faible indique une liaison plus étroite avec l’enzyme et un inhibiteur plus efficace. Dans l’ensemble, les valeurs Ki des composés étaient en corrélation avec les valeurs IC50 respectives. L’inhibition de BACE1 par les composés (2R,3R)-ptéroside C, (3S)-ptéroside D, et (2R,3R)-ptérosine C était de type mixte avec des valeurs Ki de 12,6, 16,5, et 27.6 µM, respectivement, tandis que l’inhibition par le (2R)-ptéroside B, la (2S,3R)-ptérosine C et la (2R)-ptérosine B était non compétitive avec des valeurs Ki de 23,1, 33,8 et 38,3 µM, respectivement. L’inhibition de l’AChE par le (2R)-ptéroside B, le (2R,3R)-ptéroside C, la (2R)-ptérosine B, la (2S,3R)-ptérosine C et le (3S)-ptéroside D était de type mixte avec des valeurs Ki de 4.89, 8,13, 12,1, 16,3 et 23,1 µM, respectivement, tandis que la (2R,3R)-ptérosine C était un inhibiteur de type non compétitif avec une valeur Ki de 29,6 µM. L’inhibition du BChE par la (2R,3R)-ptérosine C, le (2R,3R)-ptéroside C, le (3S)-ptéroside D et le (2R)-ptéroside B était de type mixte avec des valeurs Ki de 4,77, 9,62, 19.7, et 22,6 µM, respectivement, tandis que la (2S,3R)-ptérosine C et la (2R)-ptérosine B étaient des inhibiteurs non compétitifs avec des valeurs Ki de 29,9 et 53,5 µM, respectivement. Ainsi, ces résultats ont suggéré que des dérivés spécifiques de la ptérosine pourraient être des inhibiteurs efficaces de la BACE1, de l’AChE et de la BChE.
Simulations de docking moléculaire pour BACE1, AChE, et BChE
Plusieurs structures cristallines sont disponibles pour BACE1 et les cholinestérases. Nous avons sélectionné les PDB humaines en fonction des structures de type sauvage, des ligands cocristallisés et des résolutions des structures. Les structures cristallines aux rayons X de la BACE1 complexée avec QUD (code PDB : 2WJO, résolution : 2,5 Å)33, de l’AChE complexée avec E2020 (code PDB : 4EY7, résolution : 2,35 Å)34, et de la BChE complexée avec 3F9 (code PDB : 4TPK, résolution : 2,70 Å)35 ont été sélectionnées pour l’arrimage. Initialement, QUD, E2020 et 3F9 ont été extraits des structures cristallines et redockés dans les sites actifs de la BACE1, de l’AChE et de la BChE, respectivement. Par la suite, le (2R,3R)-ptéroside C, le (3S)-ptéroside D, le (2R,3R)-ptérosine C, le (2R)-ptéroside B, le (2S,3R)-ptérosine C et le (2R)-ptérosine B avec le mécanisme d’inhibition connu contre BACE1, AChE et BChE ont été dockés pour déterminer leur SAR. Les résultats de l’arrimage sont résumés dans le tableau 3. Le SAR des inhibiteurs de BACE1, AChE et BChE sélectionnés, de type mixte et non compétitifs, a permis d’évaluer une nouvelle inhibition des enzymes par les ptérosines, dirigée vers le site de liaison.
Notre mode de docking de E2020 était cohérent avec le mode de liaison déterminé expérimentalement précédemment rapporté avec l’AChE humaine recombinante (rhAChE) (Informations supplémentaires 3)34. La déviation quadratique moyenne (RMSD) entre le cristal et les conformations d’arrimage de E2020 était de 0,54 Å, ce qui suggère la fiabilité de notre configuration d’arrimage pour reproduire le mode de liaison expérimental. En outre, le mode d’ancrage de E2020 a conduit à une interaction similaire à celle de rhAChE-E2020. Dans notre étude, les molécules d’eau ont été retirées de la structure cristalline pendant l’amarrage ; par conséquent, les interactions médiées par l’eau n’ont pas été analysées dans la présente étude. De même, les modes d’ancrage de QUD et 3F9 étaient cohérents avec les données expérimentales disponibles pour BACE1 33 et BChE35, respectivement (Informations supplémentaires 3). Les RMSD entre le cristal et les conformations ancrées de QUD et 3F9 étaient de 0,46 et 0,60 Å, respectivement. En outre, les sites de liaison des inhibiteurs de la ptérosine étaient en accord avec une étude d’arrimage précédente qui impliquait la BACE1, l’AChE et le BChE38. Cependant, l’étude a utilisé l’AChE de Tetronarce californica (code PDB : 1ACJ), qui contient des numéros de résidus légèrement différents de ceux de l’AChE humaine en raison de variations dans leurs séquences.
Docking de BACE1
Selon le type d’inhibition et l’activité, le (2R,3R)-ptéroside C et le (2R)-ptéroside B ont été sélectionnés pour démontrer les modes docking des inhibiteurs de BACE1 de type mixte et non compétitifs, respectivement. Les figures 3a et b montrent les modèles d’ancrage du (2R,3R)-ptéroside C et du (2R)-ptéroside B, respectivement. Les interactions des composés dockés à l’intérieur du site actif de BACE1 sont présentées dans la figure 4.

Modèles de docking moléculaire pour a l’inhibiteur de type mixte de la BACE1 (2R,3R)-ptéroside C (couleur bleue) et b l’inhibiteur non compétitif de la BACE1 (2R)-ptéroside B (couleur brune). Les positions d’ancrage sont superposées à la structure cristalline de QUD (couleur jaune) (code PDB : 2WJO). La BACE1, les résidus du site actif et les composés sont représentés par des modèles en ruban, en ligne et en bâton, respectivement. Les couleurs des lignes pointillées expliquent les types de diverses interactions : interactions de liaison hydrogène (vert), interactions hydrophobes (rose) et interactions π-sigma (violet). BACE1 enzyme de clivage de la protéine précurseur de l’amyloïde β-site 1

Diagramme d’interaction du ligand de a QUD, b (2R,3R)-ptéroside C, c (3S)-ptéroside D, d (2R,3R)-ptérosine C, e (2R)-ptéroside B, f (2S,3R)-ptérosine C, et g (2R)-ptérosine B dans le site actif de la BACE1. Les couleurs des lignes pointillées expliquent les types de diverses interactions : interactions de liaison hydrogène (vert), interactions hydrophobes (rose) et interactions π-sigma (violet). BACE1 enzyme de clivage de la protéine précurseur d’amyloïde à site β 1
La pose docked de QUD a présenté une énergie de liaison (B.E.) de -7,59 kcal/mol. Comme le montre la figure 4a, le groupe NH2 sur le cycle quinazoline du ligand a montré deux liaisons hydrogène avec les groupes CO de Asp32 et Asp228 à des distances de 1,86 et 2,17 Å, respectivement. Une troisième liaison hydrogène a été observée entre l’autre groupe NH du ligand et le groupe CO de Gly230 à une distance de 2,16 Å. Leu30, Tyr71, Phe108, et Val332 ont médié les interactions hydrophobes. Les figures 4b-d montrent les positions dockées du (2R,3R)-ptéroside C, du (3S)-ptéroside D et de la (2R,3R)-ptérosine C (inhibiteurs de type mixte de la BACE1), respectivement. Ils ont été positionnés dans la poche de liaison bordée par Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 et Thr231. Selon leurs niveaux d’activité, le (2R,3R)-ptéroside C (IC50 = 9,74 µM), le (3S)-ptéroside D (IC50 = 10,7 µM) et la (2R,3R)-ptérosine C (IC50 = 26,2 µM) ont présenté une B.E. de -7,27, -6,93 et -4,84 kcal/mol, respectivement. Le (2R,3R)-Ptéroside C a présenté une puissance plus élevée que la (2R,3R)-ptérosine C en raison de l’existence d’un groupe supplémentaire 2-hydroxyméthyl-tétrahydro-pyran-3,4,5-triol, qui a formé trois liaisons hydrogène (Fig. 4b). Le groupe OH de l’hydroxyméthyle et le groupe 3-OH du cycle tétrahydro-pyrane-triol ont montré deux liaisons hydrogène avec le groupe CO de l’Asp228 à des distances de 2,63 et 2,12 Å, respectivement. De plus, le groupe 3-OH a montré une liaison hydrogène avec Thr231 à une distance de 1,89 Å. Dans le cas du (3S)-ptéroside D (Fig. 4c), la présence du groupe 2,2-diméthyle sur le cycle indanone a légèrement modifié les interactions de liaison par rapport au (2R,3R)-ptéroside C. Le groupe 3-OH du cycle tétrahydro-pyranne-triol a montré une seule liaison hydrogène avec Asp228 à une distance de 1,93 Å. Cependant, le groupe 3-OH du cycle indanone a présenté une liaison hydrogène supplémentaire avec Ile126 à une distance de 2,34 Å. Ces interactions ont légèrement diminué l’activité du (3S)-ptéroside D par rapport au (2R,3R)-ptéroside C. Les interactions d’amarrage de la (2R,3R)-ptérosine C présentées sur la figure 4d montrent que le groupe 3-OH du cycle indanone est lié à Ser36 et Asn37 par deux liaisons hydrogène à des distances de 1,83 et 2,07 Å, respectivement. Ala39, Val69, Trp76, Ile118, et Arg128 ont été impliqués dans des interactions hydrophobes, tandis que Val69 a présenté une interaction π-sigma.
Les poses dockées du (2R)-ptéroside B, de la (2S,3R)-ptérosine C, et de la (2R)-ptérosine B (inhibiteurs non compétitifs de BACE1) sont présentées dans la figure 4e-g, respectivement. Ils se sont logés dans la cavité délimitée par Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 et Ile126. Conformément à leurs valeurs d’activité, le (2R)-ptéroside B (IC50 = 18,0 µM), la (2S,3R)-ptérosine C (IC50 = 23,1 µM) et la (2R)-ptérosine B (IC50 = 29,6 µM) ont présenté une B.E. de -6,16, -5,07 et -4,64 kcal/mol, respectivement. Comme le montre la figure 4e, le (2R)-ptéroside B a montré une activité plus élevée que la (2S,3R)-ptérosine C et la (2R)-ptérosine B en raison de la présence d’un groupe supplémentaire 2-hydroxyméthyl-tétrahydro-pyran-3,4,5-triol, qui a montré quatre interactions de liaison hydrogène. Deux liaisons hydrogène ont été observées entre le groupe 4-OH du cycle tétrahydro-pyran-triol et les groupes NH et CO de l’Asn37 à des distances de 2,48 et 2,19 Å, respectivement. De plus, les groupes 3- et 5-OH ont montré deux liaisons hydrogène supplémentaires avec Ile126 et Trp76 à des distances de 2,19 et 2,46 Å, respectivement. La (2S,3R)-Ptérosine C a montré une activité légèrement meilleure que la (2R)-ptérosine B en raison de la présence d’un groupe OH supplémentaire en position 3 du cycle indanone, qui a formé une liaison hydrogène avec Lys107 à une distance de 2,12 Å (Fig. 4f). Les autres interactions étaient similaires à celles de la (2R)-ptérosine B. Comme le montre la figure 4g, la (2R)-ptérosine B a montré des interactions hydrophobes avec Val69, Tyr76 et Phe108.
Docking de l’AChE
Le (2R)-Ptéroside B et la (2R,3R)-ptérosine C ont été choisis comme représentants pour démontrer les modes de docking des inhibiteurs de l’AChE de type mixte et non compétitif, respectivement, en raison de leurs activités et du type d’inhibition de l’AChE. Les figures 5a et b illustrent les modèles d’ancrage du (2R)-ptéroside B et de la (2R,3R)-ptérosine C, respectivement. Les interactions des composés dockés à l’intérieur du site actif de l’AChE sont affichées dans la Fig. 6.

Modèles de docking moléculaire pour a l’inhibiteur de type mixte de l’AChE (2R)-ptéroside B (couleur bleue) et b l’inhibiteur non compétitif de l’AChE (2R,3R)-ptérosine C (couleur brune). Les poses sont superposées à la structure cristalline aux rayons X de E2020 (couleur jaune) (code PDB : 4EY7). L’AChE, les résidus du site actif et les composés sont représentés par des modèles en ruban, en ligne et en bâton, respectivement. Les couleurs des lignes pointillées expliquent les types de diverses interactions : interactions de liaison hydrogène (vert) et interactions hydrophobes (rose). AChE acétylcholinestérase

Diagramme d’interaction du ligand de a E2020, b (2R)-ptéroside B, c (2R,3R)-ptéroside C, d (2S,3R)-ptérosine C, e (2R)-ptérosine B, f (3S)-ptéroside D et g (2R,3R)-ptérosine C dans le site actif de l’AChE. Les couleurs des lignes pointillées expliquent les types de diverses interactions : interactions de liaison hydrogène (vert), interactions hydrophobes (rose) et interactions π-sigma (violet). AChE acétylcholinestérase
La pose dockée de E2020 a démontré une B.E. de -10,28 kcal/mol. Comme illustré dans la figure 6a, le groupe CO du cycle indanone a formé une liaison hydrogène avec le groupe NH de Phe295 à une distance de 1,70 Å. Trp286 et Tyr341 ont été impliqués dans des interactions π-sigma, tandis que Trp86, Trp286, Tyr337, Phe338 et Tyr341 ont médié des interactions hydrophobes. Les figures 6b-f montrent les positions dockées du (2R)-ptéroside B, du (2R,3R)-ptéroside C, du (2S,3R)-ptéroside C, du (2R)-ptéroside B et du (3S)-ptéroside D (inhibiteurs de l’AChE de type mixte), respectivement. Ils ont été logés dans le site actif entouré de Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 et Tyr341. Conformément à leurs valeurs d’activité, le (2R)-ptéroside B (IC50 = 2,55 µM), le (2R,3R)-ptéroside C (IC50 = 3,77 µM), la (2S,3R)-ptérosine C (IC50 = 12.8 µM), la (2R)-ptérosine B (IC50 = 16,2 µM) et le (3S)-ptéroside D (IC50 = 27,4 µM) ont présenté une B.E. de -7,90, -7,49, -6,03, -5,76 et -4,91 kcal/mol, respectivement. Le (2R)-Ptéroside B a démontré une puissance plus élevée que la (2S,3R)-ptérosine C et la (2R)-ptérosine B en raison de la présence d’un groupe supplémentaire 2-hydroxyméthyl-tétrahydro-pyran-3,4,5-triol, qui a établi trois interactions de liaison hydrogène (Fig. 6b). Le groupe OH de l’hydroxyméthyle et le groupe 3-OH du cycle tétrahydro-pyranne-triol ont présenté des liaisons hydrogène avec le groupe CO de Asn87 à des distances de 2,23 et 2,28 Å, respectivement. De plus, le groupe 4-OH a formé une liaison hydrogène avec le groupe CO de Trp86 à une distance de 2,13 Å. Dans le cas du (2R,3R)-ptéroside C (Fig. 6c), le groupe méthyle en position 2 du cycle indanone n’a pas montré d’interaction hydrophobe avec Tyr341 et a donc présenté une activité comparativement plus faible que le (2R)-ptéroside B. Cependant, Tyr341 a maintenu l’interaction hydrophobe avec l’autre partie du cycle indanone comme le montre celui du (2R)-ptéroside B. L’activité plus élevée de la (2S,3R)-ptérosine C que de la (2R)-ptérosine B a été attribuée à l’existence d’un groupe OH supplémentaire en position-3 du cycle indanone, qui a formé une liaison hydrogène avec Phe295 à une distance de 1,82 Å (Fig. 6d). Les autres interactions étaient comparables aux interactions de la (2R)-ptérosine B. Comme le montre la figure 6e, le groupe OH du groupe hydroxyéthyle en position 6 du cycle indanone a formé une liaison hydrogène avec Tyr124 à une distance de 2,48 Å. Tyr124, Trp286, Tyr337, Phe338 et Tyr341 ont contribué aux interactions hydrophobes. Comparé au (2R)-ptéroside B et au (2R,3R)-ptéroside C, le (3S)-ptéroside D a présenté des interactions de liaison différentes en raison de la présence du groupe 2,2-diméthyle sur le cycle indanone (Fig. 6f). Le groupe 2,2-diméthyle a contribué de manière significative à la pose distincte du (3S)-ptéroside D. Le groupe 5-OH du cycle tétrahydro-pyran-triol a formé une liaison hydrogène avec Asp74 à une distance de 1,96 Å. Le groupe CO du cycle indanone a montré une liaison hydrogène avec Tyr72 à une distance de 2,97 Å. Ces interactions expliquent la faible activité du (3S)-ptéroside D.
La pose docked de la (2R,3R)-ptérosine C (inhibiteur non compétitif de l’AChE) est présentée dans la figure 6g. La (2R,3R)-Ptérosine C (IC50 = 23,2 µM) a démontré une B.E. de -5,01 kcal/mol. La poche de liaison de la (2R,3R)-ptérosine C comprenait Tyr72, Trp286, Ser293, Phe297 et Tyr341, avec deux interactions par liaison hydrogène. L’une des liaisons hydrogène a été formée entre le groupe 3-OH du cycle indanone et le groupe CO de Ser293 à une distance de 2,03 Å. La deuxième liaison hydrogène a été observée entre le groupe OH du groupe hydroxyéthyle présent en position 6 du cycle indanone et Tyr72 à une distance de 1,94 Å. Les résidus tels que Tyr72, Trp286, Phe297 et Tyr341 ont participé à des interactions hydrophobes.
Docking BChE
Considérant les niveaux d’activité et le type d’inhibition du BChE, le (2R,3R)-ptéroside C et la (2S,3R)-ptérosine C ont été sélectionnés pour démontrer les modes de docking des inhibiteurs du BChE de type mixte et non compétitif, respectivement. Les figures 7a et b illustrent les modèles d’ancrage du (2R,3R)-ptéroside C et de la (2S,3R)-ptérosine C, respectivement. Les interactions des composés dockés à l’intérieur du site actif du BChE sont présentées dans la Fig. 8.
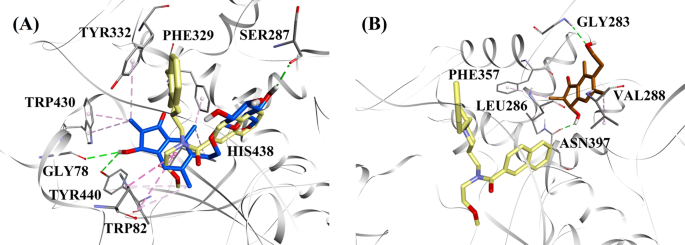
Modèles de docking moléculaire pour a l’inhibiteur de BChE de type mixte (2R,3R)-ptéroside C (couleur bleue) et b l’inhibiteur de BChE non compétitif (2S,3R)-ptérosine C (couleur brune). Les poses sont superposées à la structure cristalline aux rayons X de 3F9 (couleur jaune) (code PDB : 4TPK). Le BChE, les résidus du site actif et les composés sont représentés par des modèles en ruban, en ligne et en bâton, respectivement. Les couleurs des lignes pointillées expliquent les types de diverses interactions : interactions de liaison hydrogène (vert) et interactions hydrophobes (rose). BChE butyrylcholinestérase
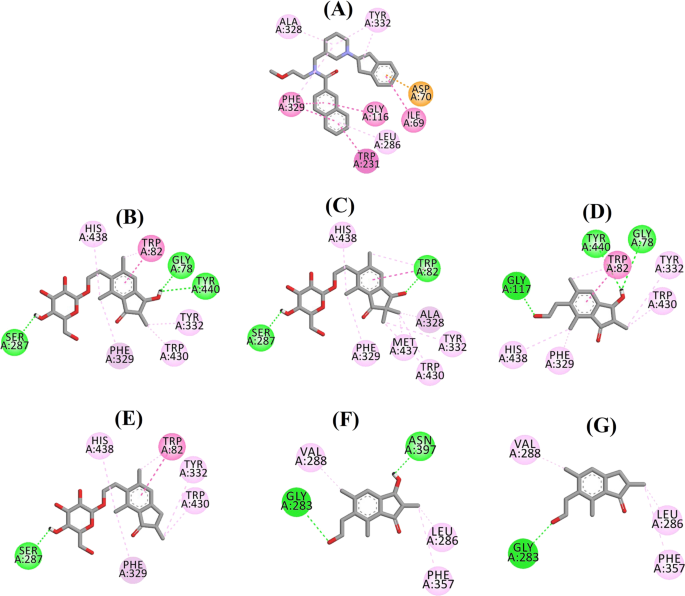
Diagramme d’interaction du ligand de a 3F9, b (2R,3R)-ptéroside C, c (3S)-ptéroside D, d (2R,3R)-ptérosine C, e (2R)-ptéroside B, f (2S,3R)-ptérosine C et g (2R)-ptérosine B dans le site actif de BChE. Les couleurs des lignes pointillées expliquent les types des diverses interactions : interactions de liaison hydrogène (vert), interactions hydrophobes (rose) et interactions π-anion (doré). BChE butyrylcholinestérase
La pose dockée de 3F9 a montré une B.E. de -8,49 kcal/mol. Comme le montre la figure 8a, les interactions hydrophobes étaient principalement responsables de la liaison du ligand. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329, et Tyr332 sont responsables des interactions hydrophobes, tandis que Asp70 a démontré une interaction π-anion. Les figures 8b-e illustrent les poses dockées du (2R,3R)-ptéroside C, du (3S)-ptéroside D, de la (2R,3R)-ptérosine C et du (2R)-ptéroside B (inhibiteurs de BChE de type mixte), respectivement. Leur poche de liaison était composée de Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 et Tyr440. Conformément à leurs niveaux d’activité, le (2R,3R)-ptéroside C (IC50 = 5,29 µM), le (3S)-ptéroside D (IC50 = 19,3 µM), la (2R,3R)-ptérosine C (IC50 = 20,3 µM) et le (2R)-ptéroside B (IC50 = 62,0 µM) ont démontré une B.E. de -7,23, -6,59, -6,52, et -4,38 kcal/mol, respectivement. Comme le montre la figure 8b, le groupe 3-OH du cycle indanone du (2R,3R)-ptéroside C a présenté deux liaisons hydrogène avec Gly78 et Tyr440 à des distances de 2,87 et 2,89 Å, respectivement. Dans le cas du (3S)-ptéroside D, le groupe 2,2-diméthyle du cycle indanone a affecté les interactions de liaison du groupe 3-OH (Fig. 8c). Le groupe 3-OH n’a formé qu’une seule liaison hydrogène avec Trp82 à une distance de 2,94 Å, ce qui a entraîné une activité comparativement plus faible du (3S)-ptéroside D que du (2R,3R)-ptéroside C. Comme le montre la figure 8d, la (2R,3R)-ptérosine C n’a pas réussi à produire une liaison hydrogène avec Ser287 en raison de l’absence d’un groupe 2-hydroxyméthyl-tétrahydro-pyran-3,4,5-triol. Par conséquent, il a donné une activité plus faible que le (2R,3R)-ptéroside C. L’absence du groupe OH en position 3 du cycle indanone s’est avérée être responsable de la très faible activité du (2R)-ptéroside B (Fig. 8e), qui n’a pas réussi à former des liaisons hydrogène avec Gly78 et Tyr440 comme le (2R,3R)-ptéroside C.
Les positions ancrées de la (2S,3R)-ptérosine C et de la (2R)-ptérosine B (inhibiteurs non compétitifs de BChE) sont présentées dans les figures 8f, g, respectivement. Ces poses amarrées étaient contenues dans la cavité délimitée par Gly283, Leu286, Val288, Phe357 et Asn397. Selon leurs niveaux d’activité, la (2S,3R)-ptérosine C (IC50 = 44,3 µM) et la (2R)-ptérosine B (IC50 = 48,1 µM) ont présenté un B.E. de -5,40 et -5,06 kcal/mol, respectivement. La (2S,3 R)-Ptérosine C était plus puissante que la (2R)-ptérosine B en raison de la présence d’un groupe OH supplémentaire en position 3 du cycle indanone, qui a formé une liaison hydrogène avec le groupe CO de l’Asn397 à une distance de 2,05 Å (Fig. 8f). D’autres interactions se sont avérées similaires à celles de la (2R)-ptérosine B. Comme le montre la figure 8g, le groupe OH du groupe hydroxyéthyle en position 6 du cycle indanone a formé une liaison hydrogène avec Gly283 à une distance de 2,40 Å. Les résidus Leu286, Val288, et Phe357 ont participé à des interactions hydrophobes.
Les inhibiteurs de type mixte se lient à la fois à l’enzyme libre et au complexe enzyme-substrat, ce qui indique que ces composés peuvent se lier au site catalytique de chaque enzyme correspondante. Les inhibiteurs non compétitifs se lient au site allostérique de l’enzyme libre ou du complexe enzyme-substrat. Une étude récente a suggéré que les inhibiteurs compétitifs, mixtes et non compétitifs occupent différents sites dans les poches de liaison de la BACE1, de l’AChE et de la BChE38. Lors du docking pour l’évaluation du mécanisme inhibiteur des dérivés de la ptérosine, les sites de liaison des composés ont été définis en fonction de leur type d’inhibition. Les résultats du docking ont indiqué que les sites de liaison des inhibiteurs de type mixte et non compétitifs pour BACE1, AChE et BChE se chevauchent partiellement à chaque site actif correspondant et étaient cohérents avec un rapport précédent38.
Perméabilité de la BHE
PAMPA-BBB, un test de perméabilité membranaire artificielle in vitro pour la BHE, est l’un des outils de criblage physico-chimique les plus fiables dans la découverte précoce de médicaments ciblant le SNC40. Le système PAMPA-BBB modélise la diffusion passive transcellulaire de substances chimiques à travers la BHE et mesure les mécanismes de transport strictement passif via une membrane lipidique artificielle sur la perméabilité effective (Pe, cm/s). Sur la base du modèle établi pour la prédiction de la perméabilité de la BHE, les composés ont été classés en (i) « SNC+ » (forte perméabilité prédite de la BHE) ; Pe (10-6 cm/s) > 4,00, (ii) « SNC- » (faible perméabilité prédite de la BHE) ; Pe (10-6 cm/s) < 2,00, et (iii) « SNC+/- » (perméabilité incertaine de la BHE) ; Pe (10-6 cm/s) de 4,00 à 2,00. En conséquence, la (2R)-ptérosine B, la (2S)-ptérosine P et la (2S)-ptérosine A ont présenté une perméation élevée de la BHE avec des valeurs Pe de 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s et 6,26 × 10-6 cm/s, respectivement (Tableau 4, Informations supplémentaires 4). La valeur Pe de la (2R)-ptérosine B était 1,7 fois plus élevée que celle du médicament du SNC, le vérapamil (Pe = 34,6 × 10-6 cm/s), qui a été utilisé comme contrôle positif. La (2S,3R)-Ptérosine C et la (2R,3R)-ptérosine C ont montré une perméabilité incertaine de la BHE avec des valeurs de Pe de 2,34 et 1,98, respectivement. Le (2R,3R)-Ptéroside C, le (3S)-ptéroside D et le (2R)-ptéroside B, qui ont montré les plus puissantes activités d’inhibition de la BACE1 et de la cholinestérase parmi les dérivés de la ptérosine testés, ont présenté une très faible perméabilité à la BHE. L’existence du groupe 2-hydroxyméthyl-tétrahydro-pyran-3,4,5-triol comme dans les dérivés de la ptérosine a entraîné une diminution remarquable de la perméabilité de la BHE. Par rapport à la (2R)-ptérosine B, la présence supplémentaire du groupe OH en position 3 du cycle indanone comme dans la ptérosine C, du groupe hydroxyméthyle en position 2 du cycle indanone comme dans la (2S)-ptérosine A ou du groupe hydroxyméthyle en position 5 du cycle indanone comme dans la (2S)-ptérosine P a également réduit de manière significative la perméabilité de la BHE. Compte tenu d’une perméabilité de la BHE exceptionnellement élevée et de l’inhibition significative de la BACE1, de l’AChE et de la BChE, la (2R)-ptérosine B pourrait avoir le potentiel de présenter une forte activité anti-AD.
Effets de la (2R)-ptérosine B et du (2R,3R)-ptéroside C sur la sécrétion des peptides Aβ par les cellules neuronales
Pour étudier la fonction de la (2R)-ptérosine B et du (2R,3R)-ptéroside C dans la diminution de l’excrétion d’Aβ par les cellules neuronales, nous avons utilisé une lignée cellulaire de neuroblastome murin qui surexprime de façon stable l’APPswe humaine. Cette lignée cellulaire est un modèle cellulaire de la MA caractérisé par la sécrétion excessive d’Aβ40 et d’Aβ42. Les oligomères amyloïdes toxiques sont formés à partir des deux isoformes de peptide Aβ de longueurs différentes. L’Aβ40 est l’isoforme d’Aβ la plus abondante dans le cerveau41, tandis que l’Aβ42 augmente significativement avec certaines formes de MA42. L’ELISA sandwich de l’Aβ40 a montré que la (2R)-ptérosine B réduisait significativement la quantité de peptide Aβ40 sécrétée par les cellules de neuroblastome dans les milieux jusqu’à 50 % à 500 μM (P < 0,01) (figure 9a). De même, la sécrétion du peptide Aβ42 par les cellules de neuroblastome a significativement diminué en présence de 500 µM de (2R,3R)-ptéroside C (P < 0,05) (Fig. 9b). En conclusion, la (2R)-ptérosine B et le (2R,3R)-ptéroside C ont significativement diminué la sécrétion des peptides Aβ des cellules de neuroblastome à une concentration de 500 μM.

a Effet de la (2R)-ptérosine B sur la sécrétion des peptides Aβ. Des cellules de neuroblastome sécrétant de l’APPswe ont été traitées avec 12, 60, 250 ou 500 µM de (2R)-ptérosine B pendant 24 h, et les milieux conditionnés ont été recueillis en présence d’un inhibiteur de protéase. La β-SI (10 µM) a été utilisée comme contrôle positif. Le contrôle négatif comprenait des cellules cultivées en l’absence des composés testés. L’analyse quantitative de l’Aβ40 et de l’Aβ42 sécrétés dans les milieux conditionnés a été réalisée par ELISA sandwich. Les moyennes ± SD de trois expériences indépendantes sont présentées. Les peptides Aβ40 sécrétés ont significativement diminué en présence de 500 µM de (2R)-ptérosine B. CON contrôle négatif, β-SI β-inhibiteur de la sécrétase III, Aβ β-amyloïde. *P < 0,05, **P < 0,01. b Effet du (2R,3R)-ptéroside C sur la sécrétion des peptides Aβ. Les cellules de neuroblastome sécrétant de l’APPswe ont été traitées avec 12, 60, 125, 250 ou 500 µM de (2R,3R)-ptéroside C pendant 24 h, et les milieux conditionnés ont été recueillis en présence d’un inhibiteur de protéase. La β-SI (10 µM) a été utilisée comme contrôle positif. Le contrôle négatif comprenait des cellules cultivées en l’absence des composés testés. L’analyse quantitative de l’Aβ40 et de l’Aβ42 sécrétés dans les milieux conditionnés a été réalisée à l’aide d’un ELISA sandwich. Les moyennes ± SD de trois expériences indépendantes sont présentées. Le peptide Aβ42 sécrété a significativement diminué en présence de 500 µM de (2R,3R)-ptéroside C. CON contrôle négatif, β-SI β-inhibiteur de la sécrétase III, Aβ β-amyloïde. *P < 0.05
Cytotoxicité des dérivés de la ptérosine basée sur le test MTT43
Dans l’ensemble, les dérivés de la ptérosine ont montré une cytotoxicité négligeable contre diverses lignées cellulaires normales et cancéreuses, telles que le neuroblastome humain SH-SY5Y, les cellules gliales de rat C6, les fibroblastes embryonnaires de souris NIH3T3 et le mélanome de souris B16F10 avec des valeurs de LD50 supérieures à 0.5 mM (Informations supplémentaires 5). En particulier, la (2R)-ptérosine B, la (3R)-ptérosine D, la (2S)-ptérosine P, le (2S)-ptéroside A, le (2R)-ptéroside B et le (2R,3R)-ptéroside C n’ont montré aucune cytotoxicité contre les lignées cellulaires testées avec des valeurs de LD50 supérieures à 5 mM. De manière intrigante, plusieurs ptérosines ont montré des effets antiprolifératifs relatifs contre les cellules neuronales SH-SY5Y par rapport aux cellules gliales C6 d’origine mésenchymateuse. Les présents résultats étaient conformes à un rapport précédent qui suggérait que les dérivés de la ptérosine ne sont pas toxiques pour les humains26.