
Le contexte environnemental de l’adaptation
Nous avons déjà parlé à plusieurs reprises de la façon dont les espèces évoluent en réponse à une sorte de pression environnementale, qui favorise (ou défavorise) certains traits au sein de cette espèce. Au fil du temps, cela entraîne des changements dans les fréquences des traits de l’espèce et modifie le phénotype moyen global de cette espèce (parfois lentement, parfois rapidement).
Si nous parlons généralement de l’environnement en termes de conditions abiotiques telles que la température ou le climat, les facteurs biotiques sont tout aussi importants : c’est-à-dire les parties de l’environnement qui sont elles-mêmes également vivantes. De ce fait, l’évolution d’une espèce peut avoir de profondes répercussions sur les autres espèces liées au sein de l’écosystème. Ainsi, l’évolution d’une espèce est intrinsèquement liée à l’évolution d’autres espèces pertinentes au sein de l’écosystème : souvent, ces voies d’évolution connectées s’affrontent lorsque chacune change. Voyons quelques exemples différents de la façon dont l’évolution d’une espèce peut avoir un impact sur l’évolution d’une autre.
Coévolution prédateur-proie
L’une des façons les plus évidentes dont l’évolution de deux espèces différentes peut interagir est dans les relations entre prédateurs et proies. Naturellement, les espèces proies évoluent pour pouvoir se défendre des prédateurs de diverses manières, comme la crypse (par exemple le camouflage), la toxicité ou les changements de comportement (comme le nocturnisme ou le rassemblement en groupe). À l’inverse, les prédateurs évolueront vers des méthodes nouvelles et améliorées pour détecter et chasser leurs proies, comme des sens accrus, du venin et de la furtivité (grâce à des pieds à rembourrage souple, par exemple).
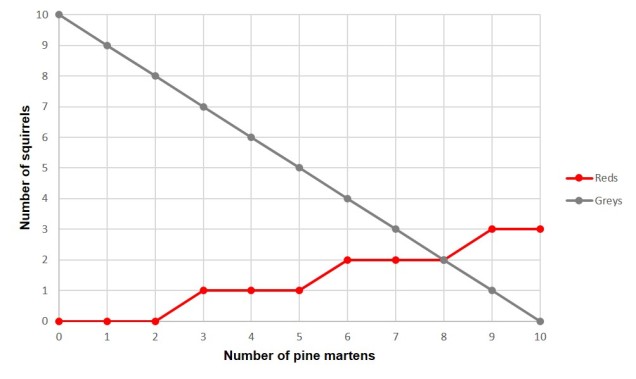
Il existe des millions d’exemples possibles de coévolution prédateur-proie qui pourraient servir d’exemples ici, basés sur la volonté continuelle d’une espèce de prendre le dessus sur l’autre. Mais l’un d’eux qui me vient à l’esprit est celui d’une créature dont j’ai appris l’existence lors de mes vacances en Scandinavie : la martre des pins, et la façon dont elle affecte les écureuils.

La martre des pins est une espèce de la famille des mustélidés, avec la loutre, la belette, la stoat et le glouton. Comme de nombreux mustélidés, ce sont des mammifères carnivores qui se nourrissent d’une variété de proies différentes comme les rongeurs, les petits oiseaux et les insectes. L’une des espèces les plus abondantes dont elles s’attaquent est l’écureuil : les écureuils roux et les écureuils gris sont tous deux des aliments potentiels pour la mignonne mais sauvage martre des pins.
Cependant, dans l’aire de répartition des martres des pins (à travers une grande partie de l’Europe), les écureuils roux sont l’espèce indigène et les écureuils gris sont invasifs, originaires d’Amérique du Nord. En raison de la relation durable entre l’écureuil roux et la martre des pins, ces deux espèces ont évolué ensemble, notamment grâce à l’écureuil roux qui a adopté un mode de vie essentiellement arboricole et qui évite le sol autant que possible. Les écureuils gris, en revanche, n’ont pas eu l’évolution nécessaire pour apprendre cette leçon et constituent une nourriture facile pour une martre des pins intelligente. Ainsi, dans les régions où les martres des pins ont été conservées ou réintroduites, elles contrôlent activement la population envahissante d’écureuils gris, ce qui a pour effet de stimuler la population indigène d’écureuils roux en réduisant la concurrence. Le lien coévolutif entre les écureuils roux et les martres des pins est essentiel pour lutter contre les espèces invasives.

Coévolution hôte-parasite
Dans une veine similaire à la coévolution des prédateurs et des proies, les espèces pathogènes et leurs hôtes malheureux subissent également une sorte de « course aux armements ». Les parasites doivent sans cesse évoluer vers de nouveaux moyens d’infecter et de transmettre à leurs hôtes, tandis que ces derniers développent de nouvelles méthodes pour résister et éviter l’espèce infectante. Cette bataille en spirale des forces évolutives est appelée « hypothèse de la reine rouge », formulée en 1973 par Leigh Van Valen et utilisée pour décrire de nombreuses autres formes de coévolution. Le nom vient de l’ouvrage de Lewis Carroll, Through the Looking Glass, et d’une citation en particulier:
‘Now, here, you see, it takes all the running you can do, to keep in the same place’.
La citation fait référence à la façon dont les espèces doivent continuellement s’adapter et répondre à l’évolution d’autres espèces pour simplement continuer à exister et éviter l’extinction. Les espèces qui restent statiques et cessent d’évoluer vont inévitablement s’éteindre à mesure que le monde qui les entoure change.
Mimétisme
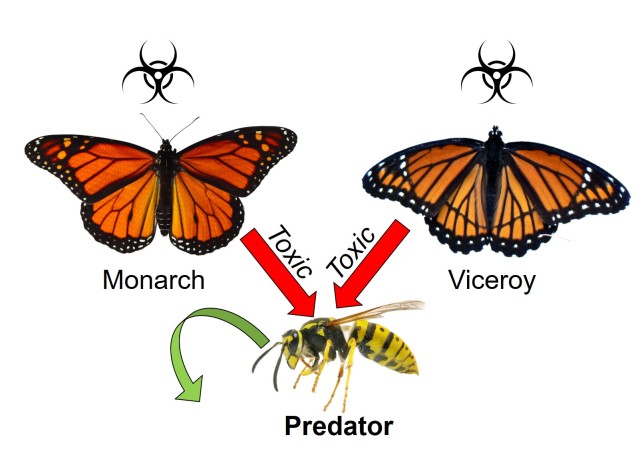
Plusieurs autres mécanismes étranges et uniques de coévolution existent dans la nature. L’un d’eux est le mimétisme, le processus par lequel une espèce tente de ressembler à une autre pour se protéger. Le groupe le plus emblématique connu pour cela est celui des papillons : de nombreuses espèces, bien qu’elles puissent être très différentes sur le plan de l’évolution, partagent des motifs de coloration et des formes corporelles similaires en tant que mimétiques. Selon la nature de la copie, le mimétisme peut être classé en deux grandes catégories. Dans les deux cas, l’espèce « de référence » initiale est toxique ou désagréable pour les prédateurs et utilise un type de signal coloré pour le faire savoir : pensez au jaune vif des abeilles et des guêpes ou au rouge des coccinelles. Là où les deux catégories changent, c’est dans la nature de l’espèce » mimétique « .
Mimétisme müllerien
Si l’espèce mimétique est également toxique ou peu appétissante, on parle de mimétisme müllerien (d’après Johann Friedrich Theodor Müller). En partageant les mêmes motifs de coloration et en étant toutes deux toxiques, les deux espèces mimétiques augmentent le potentiel d’apprentissage du signal par les prédateurs. Si un prédateur mange l’une ou l’autre des espèces, il associera ce motif de coloration à la toxicité et aucune des deux espèces ne sera plus susceptible d’être attaquée à l’avenir. En ce sens, il s’agit d’une relation de coévolution coopérative entre les deux espèces physiquement similaires.
Mimétisme batesien
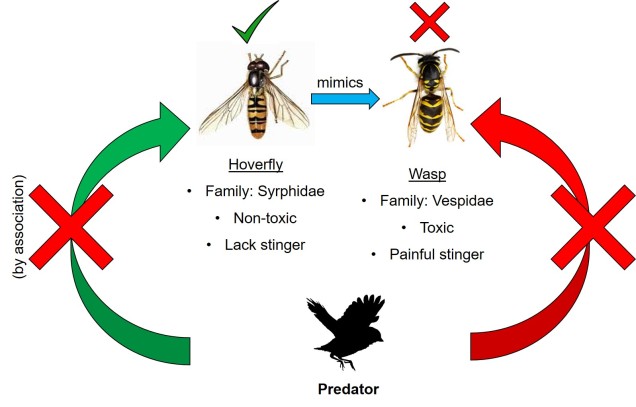
A l’inverse, le mimétique pourrait ne pas être réellement toxique ou peu appétissant, et simplement copier une espèce toxique. C’est ce que l’on appelle le mimétisme batesien (d’après Henry Walter Bates), et cela implique qu’une espèce mimétique compte sur l’association de la couleur et de la toxicité pour avoir été apprise par les prédateurs à travers l’espèce » de référence « . Bien que l’espèce mimétique ne soit pas toxique, elle s’appuie essentiellement sur le travail évolutif difficile qui a déjà été effectué par l’espèce réellement toxique. Dans ce cas, la relation coévolutive est plus parasitaire, car le mimic bénéficie de la » référence » mais la faveur n’est pas retournée.
Coevolution des espèces et importance des interactions entre espèces
Il existe d’innombrables autres interactions entre espèces qui pourraient conduire à des relations coévolutives dans la nature. Il peut s’agir de diverses formes de symbiose, ou de la réponse de différentes espèces aux ingénieurs de l’écosystème : c’est-à-dire des espèces qui peuvent changer et façonner l’environnement qui les entoure (comme les coraux dans les systèmes récifaux). Pour comprendre comment une espèce évolue dans son environnement, il faut donc tenir compte de la façon dont de nombreuses autres espèces locales évoluent également et réagissent à leur manière.