
Isolering och karakterisering av pterosinderivat från P. aquilinum
Pterosinderivat isolerades från hela växter av P. aquilinum. För undersökning av de fytokemiska beståndsdelarna från brakörteln delades ett vattenextrakt successivt med etylacetat (EtOAc) och n-butanol (n-BuOH) (fig. 1). Upprepad kolonnkromatografi av den EtOAc-lösliga fraktionen resulterade i isolering av nio derivat, inklusive (2R)-pterosin B (1), pterosin Z (2), (2S)-pterosin P (3), (3R)-pterosin D (4), (2S)-pterosin A (5), (2S,3R)-pterosin C (6), (2R,3R)-pterosin C (7), (2R)-pterosid B (8) och pterosid Z (9), med en renhet på mer än 97 % (Fig. 2). Den upprepade kolonnkromatografiska separationen av den n-BuOH-lösliga fraktionen resulterade i isolering av sex derivat, inklusive (2S)-pterosid A (10), (2S)-pterosid A2 (11), (2S,3R)-pterosid C (12), (2R,3R)-pterosid C (13), (3S)-pterosid D (14) och (2S)-pterosid P (15), med en renhet som var högre än 97 %. Föreningarnas strukturer identifierades genom analys av spektraldata, inklusive MS, 1D- och 2D-NMR (kompletterande information 1).

Strukturer av pterosinföreningar 1-15xxx
Hämmande aktivitet hos pterosinderivat mot BACE1, AChE, och BChE
För att utvärdera anti-AD-potentialen utvärderades den hämmande aktiviteten hos varje pterosinförening mot BACE1 och kolinesteraser genom respektive in vitro-inhibitionsanalyser (tabell 1). Alla testade pterosinderivat visade koncentrationsberoende hämmande aktiviteter mot BACE1 med ett intervall av IC50-värden (halvmaximal hämmande koncentration) på 9,74-94,4 μM, med undantag för (2S)-pterosin A och (2S)-pterosid P som var inaktiva vid de testade koncentrationerna, jämfört med IC50 för quercetin som användes som positiv kontroll, som var 18,8 μM. Den hämmande potensen hos de starkaste inhibitorerna var i ordningen (2R,3R)-pterosid C, (3S)-pterosid D, (2R)-pterosid B, (2S,3R)-pterosin C, (2R,3R)-pterosin C, (2S,3R)-pterosid C, (2S,3R)-pterosid C och (2R)-pterosin B med IC50-värden på 9.74, 10,7, 18,0, 23,1, 26,2, 28,9 respektive 29,6 μM. Vi testade därefter den hämmande potentialen hos pterosinderivaten mot AChE. Alla testade föreningar visade betydande AChE-hämmande aktiviteter, med IC50-värden i intervallet 2,55-110 μM, jämfört med IC50 mot AChE för berberin som användes som positiv kontroll, som var 0,39 μM. De pterosinföreningar som uppvisade den starkaste hämmande aktiviteten mot AChE var (2R)-pterosid B, (2R,3R)-pterosid C, (2S,3R)-pterosid C, (2S,3R)-pterosin C och (2R)-pterosin B med IC50-värden på 2,55, 3,77, 9,17, 12,8 respektive 16,2 μM. Slutligen testade vi den hämmande förmågan hos pterosinderivaten mot BChE. Alla testade pterosinföreningar visade hämmande aktivitet mot BChE, med IC50-värden som varierade från 5,29 till 119 μM, med undantag för (3R)-pterosin D som var inaktivt vid de testade koncentrationerna, jämfört med berberins IC50 mot BChE, som var 3,32 μM. De pterosinföreningar som uppvisade den starkaste hämmande aktiviteten mot BChE var (2R,3R)-pterosid C och pterosid Z med IC50-värden på 5,29 respektive 5,31 μM.
Kollektivt uppvisade de flesta av de testade pterosinderivaten signifikanta hämmande aktiviteter mot BACE1, AChE och BChE samtidigt. Närvaron av den extra 2-hydroximetyl-tetrahydro-pyran-3,4,5-triolgruppen som i pterosidderivaten ökade signifikant de hämmande aktiviteterna mot enzymerna. Dessutom minskade närvaron av den extra hydroximetylgruppen i position 2 i indanonringen i (2R)-pterosin B som i (2S)-pterosin A eller metylgruppen som i (3R)-pterosin D och pterosin Z de hämmande aktiviteterna mot enzymerna. Särskilt närvaron av hydroximetylgruppen vid position-5 i indanonringen som i (2S)-pterosin P minskade den hämmande aktiviteten mot BACE1.
Kinetiska parametrar för enzymhämning av pterosinderivat
I ett försök att förklara hur den enzymatiska hämmningen av pterosinderivat går till utförde vi en kinetisk analys av BACE1 och kolinesteraser för representativa hämmare (tabell 2, tilläggsinformation 2). En låg Ki (inhibitionskonstant) indikerar tätare enzymbindning och en effektivare hämmare. Överlag korrelerade föreningarnas Ki-värden med respektive IC50-värden. BACE1-hämning av föreningarna (2R,3R)-pterosid C, (3S)-pterosid D och (2R,3R)-pterosin C var av blandad typ med Ki-värden på 12,6, 16,5 och 27.6 µM, medan inhiberingen av (2R)-pterosid B, (2S,3R)-pterosin C och (2R)-pterosin B var icke-kompetitiv med Ki-värden på 23,1, 33,8 respektive 38,3 µM. AChE-hämning av (2R)-pterosid B, (2R,3R)-pterosid C, (2R)-pterosin B, (2S,3R)-pterosin C och (3S)-pterosid D var av blandad typ med Ki-värden på 4.89, 8,13, 12,1, 16,3 respektive 23,1 µM, medan (2R,3R)-pterosin C var en hämmare av icke-kompetitiv typ med ett Ki -värde på 29,6 µM. BChE-hämning av (2R,3R)-pterosin C, (2R,3R)-pterosid C, (3S)-pterosid D och (2R)-pterosid B var av blandad typ med Ki-värden på 4,77, 9,62, 19.7 respektive 22,6 µM, medan (2S,3R)-pterosin C och (2R)-pterosin B var icke-kompetitiva hämmare med Ki-värden på 29,9 respektive 53,5 µM. Dessa resultat tyder således på att specifika pterosinderivat kan vara effektiva BACE1-, AChE- och BChE-hämmare.
Molekylära dockningssimuleringar för BACE1, AChE och BChE
Flera kristallstrukturer finns tillgängliga för BACE1 och kolinesteraser. Vi valde mänskliga PDB:er baserat på vildtypstrukturer, samkristalliserade ligander och upplösningar av strukturerna. Röntgenkristallstrukturer av BACE1 komplex med QUD (PDB-kod: 2WJO, upplösning: 2,5 Å)33, AChE komplex med E2020 (PDB-kod: 4EY7, upplösning: 2,35 Å)34 och BChE komplex med 3F9 (PDB-kod: 4TPK, upplösning: 2,70 Å)35 valdes ut för dockning. Inledningsvis extraherades QUD, E2020 och 3F9 från kristallstrukturer och redockades till BACE1:s, AChE:s respektive BChE:s aktiva platser. Därefter dockades (2R,3R)-pterosid C, (3S)-pterosid D, (2R,3R)-pterosin C, (2R)-pterosid B, (2S,3R)-pterosin C och (2R)-pterosin B med den kända hämmningsmekanismen mot BACE1, AChE och BChE för att bestämma deras SAR. Dockningsresultaten sammanfattas i tabell 3. SAR för de utvalda blandade och icke-kompetitiva BACE1-, AChE- och BChE-hämmarna gjorde det möjligt att utvärdera nya bindningsplatsinriktade hämningar av enzymerna med hjälp av pterosiner.
Vårt dockningsläge för E2020 överensstämde med det experimentellt bestämda bindningsläget som tidigare rapporterats med rekombinant humant mänsklig AChE (rhAChE) (kompletterande information 3)34. RMSD-avvikelsen (root-mean-square deviation) mellan kristallkonformationen och den dockade konformationen av E2020 var 0,54 Å, vilket tyder på att vår dockningsuppsättning är tillförlitlig när det gäller att reproducera det experimentella bindningsläget. Dessutom ledde E2020:s dockade läge till en liknande interaktion som rhAChE-E2020. I vår studie avlägsnades vattenmolekyler från kristallstrukturen under dockningen; därför analyserades inte vattenmedierade interaktioner i denna studie. På samma sätt överensstämde de dockade lägena för QUD och 3F9 med tillgängliga experimentella data för BACE1 33 respektive BChE35 (kompletterande information 3). RMSD mellan kristallkonformationerna och de dockade konformationerna av QUD och 3F9 var 0,46 respektive 0,60 Å. Vidare överensstämde pterosinhämmarnas bindningsställen med en tidigare dockningsstudie som omfattade BACE1, AChE och BChE38. I studien användes dock Tetronarce californica AChE (PDB-kod: 1ACJ), som innehåller något annorlunda restnummer än mänsklig AChE på grund av variationer i deras sekvenser.
BACE1-dockning
Baserat på hämmningstyp och aktivitet valdes (2R,3R)-pterosid C och (2R)-pterosid B ut för att demonstrera de dockade lägena för blandade typer av respektive icke-kompetitiva BACE1-hämmare. I figur 3a och b visas dockningsmodellerna för (2R,3R)-pterosid C respektive (2R)-pterosid B. Interaktionerna mellan de dockade föreningarna i den aktiva platsen för BACE1 visas i figur 4.

Molekylära dockningsmodeller för a BACE1-hämmaren av blandad typ (2R,3R)-pterosid C (blå färg) och b den icke-kompetitiva BACE1-hämmaren (2R)-pterosid B (brun färg). De dockade positionerna är överlagrade på röntgenkristallstrukturen av QUD (gul färg) (PDB-kod: 2WJO). BACE1, rester på den aktiva platsen och föreningar visas med band-, linje- och stavmodeller. Färgerna på de streckade linjerna förklarar typerna av olika interaktioner: vätebindningsinteraktioner (grönt), hydrofoba interaktioner (rosa) och π-sigmainteraktioner (lila). BACE1 β-site amyloid precursor protein cleaving enzyme 1

Ligandinteraktionsdiagram för a QUD, b (2R,3R)-pterosid C, c (3S)-pterosid D, d (2R,3R)-pterosin C, e (2R)-pterosid B, f (2S,3R)-pterosin C och g (2R)-pterosin B i den aktiva platsen för BACE1. Färgerna på de streckade linjerna förklarar typerna av olika interaktioner: vätebindningsinteraktioner (grönt), hydrofoba interaktioner (rosa) och π-sigmainteraktioner (lila). BACE1 β-site amyloid precursor protein cleaving enzyme 1
Den dockade posen av QUD uppvisade en bindningsenergi (B.E.) på -7,59 kcal/mol. Som framgår av figur 4a visade NH2-gruppen på ligandens quinazolinring två vätebindningar med CO-grupperna i Asp32 och Asp228 på avstånden 1,86 respektive 2,17 Å. En tredje vätebindning observerades mellan ligandens andra NH-grupp och CO-gruppen hos Gly230 på ett avstånd av 2,16 Å. Leu30, Tyr71, Phe108 och Val332 förmedlade de hydrofoba interaktionerna. Figur 4b-d visar de dockade positionerna för (2R,3R)-pterosid C, (3S)-pterosid D och (2R,3R)-pterosin C (BACE1-hämmare av blandad typ). De placerades i den bindningsficka som kantas av Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228 och Thr231. Enligt deras aktivitetsnivåer uppvisade (2R,3R)-pterosid C (IC50 = 9,74 µM), (3S)-pterosid D (IC50 = 10,7 µM) och (2R,3R)-pterosin C (IC50 = 26,2 µM) en B.E. på -7,27, -6,93 respektive -4,84 kcal/mol. (2R,3R)-Pterosid C uppvisade en högre potens än (2R,3R)-Pterosin C på grund av förekomsten av en ytterligare 2-hydroximetyl-tetrahydro-pyran-3,4,5-triolgrupp, som bildade tre vätebindningar (fig. 4b). OH-gruppen i hydroximetyl och 3-OH-gruppen i tetrahydro-pyrantriolringen visade två vätebindningar med CO-gruppen i Asp228 på avstånden 2,63 respektive 2,12 Å. Dessutom uppvisade 3-OH-gruppen en vätebindning med Thr231 på ett avstånd av 1,89 Å. När det gäller (3S)-pterosid D (fig. 4c) förändrade närvaron av 2,2-dimetylgruppen vid indanonringen bindningsinteraktionerna något jämfört med (2R,3R)-pterosid C. 3-OH-gruppen i tetrahydropyrantriolringen uppvisade endast en enda vätebindning med Asp228 på ett avstånd av 1,93 Å. Indanonringens 3-OH-grupp uppvisade dock ytterligare en vätebindning med Ile126 på ett avstånd av 2,34 Å. Dessa interaktioner minskade något aktiviteten hos (3S)-pterosid D jämfört med (2R,3R)-pterosid C. Dockningsinteraktionerna för (2R,3R)-pterosin C som visas i figur 4d visar att indanonringens 3-OH-grupp är bunden till Ser36 och Asn37 via två vätebindningar på avstånden 1,83 respektive 2,07 Å. Ala39, Val69, Trp76, Ile118 och Arg128 var involverade i hydrofoba interaktioner, medan Val69 uppvisade en π-sigmainteraktion.
De dockade poserna för (2R)-pterosid B, (2S,3R)-pterosin C och (2R)-pterosin B (icke-kompetitiva BACE1-hämmare) visas i fig. 4e-g respektive. De dockades in i det hålrum som omges av Asn37, Val69, Tyr71, Trp76, Lys107, Phe108 och Ile126. I enlighet med deras aktivitetsvärden uppvisade (2R)-pterosid B (IC50 = 18,0 µM), (2S,3R)-pterosin C (IC50 = 23,1 µM) och (2R)-pterosin B (IC50 = 29,6 µM) en B.E. på -6,16, -5,07 respektive -4,64 kcal/mol. Som visas i figur 4e uppvisade (2R)-pterosid B högre aktivitet än (2S,3R)-pterosin C och (2R)-pterosin B på grund av närvaron av en ytterligare 2-hydroximetyl-tetrahydro-pyran-3,4,5-triolgrupp, som uppvisade fyra vätebindningsinteraktioner. Två vätebindningar observerades mellan 4-OH-gruppen i tetrahydro-pyrantriolringen och NH- och CO-grupperna i Asn37 på avstånden 2,48 respektive 2,19 Å. Vidare uppvisade 3- och 5-OH-grupperna ytterligare två vätebindningar med Ile126 och Trp76 på avstånden 2,19 respektive 2,46 Å. (2S,3R)-Pterosin C visade något bättre aktivitet än (2R)-Pterosin B på grund av närvaron av ytterligare en OH-grupp i position 3 i indanonringen, som bildade en vätebindning med Lys107 på ett avstånd av 2,12 Å (fig. 4f). De andra interaktionerna liknade dem för (2R)-pterosin B. Som visas i figur 4g visade (2R)-pterosin B hydrofoba interaktioner med Val69, Tyr76 och Phe108.
AChE-dockning
(2R)-Pterosid B och (2R,3R)-pterosin C valdes ut som representanter för att visa dockningslägena för AChE-hämmare av blandad typ respektive icke-kompetitiva AChE-hämmare, på grund av deras aktiviteter och typ av AChE-hämning. Figur 5a och b visar dockningsmodellerna för (2R)-pterosid B respektive (2R,3R)-pterosin C. Interaktionerna mellan de dockade föreningarna i AChE:s aktiva plats visas i figur 6.

Molekylära dockningsmodeller för a AChE-hämmaren av blandad typ (2R)-pterosid B (blå färg) och b den icke-kompetitiva AChE-hämmaren (2R,3R)-pterosin C (brun färg). De dockade positionerna är överlagrade på röntgenkristallstrukturen av E2020 (gul färg) (PDB-kod: 4EY7). AChE, aktiva platsrester och föreningar visas med band-, linje- och stavmodeller. Färgerna på de streckade linjerna förklarar typerna av olika interaktioner: vätebindningsinteraktioner (grönt) och hydrofoba interaktioner (rosa). AChE acetylkolinesteras

Ligandinteraktionsdiagram för a E2020, b (2R)-pterosid B, c (2R,3R)-pterosid C, d (2S,3R)-pterosin C, e (2R)-pterosin B, f (3S)-pterosid D och g (2R,3R)-pterosin C i den aktiva platsen för AChE. Färgerna på de streckade linjerna förklarar typerna av olika interaktioner: vätebindningsinteraktioner (grönt), hydrofoba interaktioner (rosa) och π-sigmainteraktioner (lila). AChE acetylkolinesteras
Den dockade ställningen av E2020 visade en B.E. på -10,28 kcal/mol. Som illustreras i fig. 6a bildade indanonringens CO-grupp en vätebindning med Phe295:s NH-grupp på ett avstånd av 1,70 Å. Trp286 och Tyr341 var involverade i π-sigmainteraktioner, medan Trp86, Trp286, Tyr337, Phe338 och Tyr341 medierade hydrofoba interaktioner. Figur 6b-f visar de dockade positionerna för (2R)-pterosid B, (2R,3R)-pterosid C, (2S,3R)-pterosin C, (2R)-pterosin B och (3S)-pterosid D (AChE-hämmare av blandad typ). De placerades i den aktiva platsen omgiven av Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338 och Tyr341. I överensstämmelse med deras aktivitetsvärden är (2R)-pterosid B (IC50 = 2,55 µM), (2R,3R)-pterosid C (IC50 = 3,77 µM), (2S,3R)-pterosin C (IC50 = 12.8 µM), (2R)-pterosin B (IC50 = 16,2 µM) och (3S)-pterosid D (IC50 = 27,4 µM) uppvisade en B.E. på -7,90, -7,49, -6,03, -5,76 respektive -4,91 kcal/mol. (2R)-Pterosid B uppvisade en högre potens än (2S,3R)-Pterosin C och (2R)-Pterosin B på grund av närvaron av en ytterligare 2-hydroximetyl-tetrahydro-pyran-3,4,5-triolgrupp, som etablerade tre vätebindningsinteraktioner (fig. 6b). OH-gruppen i hydroximetyl och 3-OH-gruppen i tetrahydro-pyran-triolringen uppvisade vätebindningar med CO-gruppen i Asn87 på avstånden 2,23 respektive 2,28 Å. Vidare bildade 4-OH-gruppen en vätebindning med CO-gruppen i Trp86 på ett avstånd av 2,13 Å. När det gäller (2R,3R)-pterosid C (fig. 6c) uppvisade metylgruppen i indanonringens position 2 ingen hydrofob interaktion med Tyr341 och uppvisade därför jämförelsevis lägre aktivitet än (2R)-pterosid B. Tyr341 bibehöll dock den hydrofoba interaktionen med den andra delen av indanonringen, vilket visades i (2R)-pterosid B. Den högre aktiviteten hos (2S,3R)-pterosin C än (2R)-pterosin B tillskrevs förekomsten av ytterligare en OH-grupp i indanonringens position-3, som bildade en vätebindning med Phe295 på ett avstånd av 1,82 Å (fig. 6d). De övriga interaktionerna var jämförbara med (2R)-pterosin B-interaktionerna. Som framgår av fig. 6e bildade OH-gruppen i hydroxyetylgruppen i indanonringens position 6 en vätebindning med Tyr124 på ett avstånd av 2,48 Å. Tyr124, Trp286, Tyr337, Phe338 och Tyr341 bidrog till de hydrofoba interaktionerna. Jämfört med (2R)-pterosid B och (2R,3R)-pterosid C uppvisade (3S)-pterosid D olikartade bindningsinteraktioner på grund av närvaron av 2,2-dimetylgruppen vid indanonringen (fig. 6f). 2,2-dimetylgruppen bidrog avsevärt till (3S)-pterosid D:s distinkta dockningsposition. 5-OH-gruppen i tetrahydro-pyrantriolringen bildade en vätebindning med Asp74 på ett avstånd av 1,96 Å. CO-gruppen i indanonringen visade en vätebindning med Tyr72 på ett avstånd av 2,97 Å. Dessa interaktioner förklarade den låga aktiviteten hos (3S)-pterosid D.
Den dockade positionen för (2R,3R)-pterosin C (icke-kompetitiv AChE-hämmare) visas i fig. 6g. (2R,3R)-Pterosin C (IC50 = 23,2 µM) uppvisade en B.E. på -5,01 kcal/mol. Bindningsfickan för (2R,3R)-pterosin C bestod av Tyr72, Trp286, Ser293, Phe297 och Tyr341, med två vätebindningsinteraktioner. En av vätebindningarna bildades mellan indanonringens 3-OH-grupp och Ser293:s CO-grupp på ett avstånd av 2,03 Å. Den andra vätebindningen observerades mellan OH-gruppen i den hydroxyetylgrupp som finns i indanonringens position 6 och Tyr72 på ett avstånd av 1,94 Å. Rester som Tyr72, Trp286, Phe297 och Tyr341 deltog i hydrofoba interaktioner.
BChE-dockning
Med tanke på aktivitetsnivåerna och typen av BChE-hämning valdes (2R,3R)-pterosid C och (2S,3R)-pterosin C ut för att demonstrera dockningslägena för BChE-hämmare av blandad typ respektive icke-kompetitiv typ. Figur 7a och b visar dockningsmodellerna för (2R,3R)-pterosid C respektive (2S,3R)-pterosin C. Interaktionerna mellan de dockade föreningarna i den aktiva platsen för BChE presenteras i figur 8.
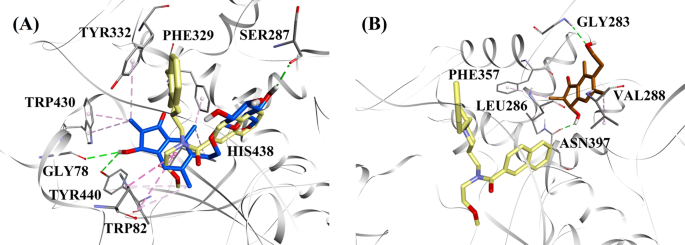
Molekylära dockningsmodeller för a BChE-hämmaren av blandad typ (2R,3R)-pterosid C (blå färg) och b den icke-kompetitiva BChE-hämmaren (2S,3R)-pterosin C (brun färg). De dockade positionerna är överlagrade på röntgenkristallstrukturen av 3F9 (gul färg) (PDB-kod: 4TPK). BChE, aktiva platsrester och föreningar visas med band-, linje- och stavmodeller. Färgerna på de streckade linjerna förklarar typerna av olika interaktioner: vätebindningsinteraktioner (grönt) och hydrofoba interaktioner (rosa). BChE butyrylkolinesteras
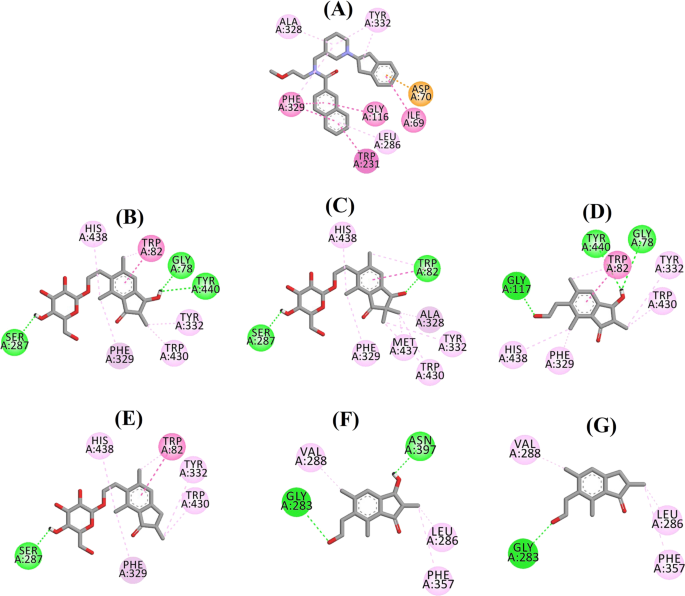
Ligandinteraktionsdiagram för a 3F9, b (2R,3R)-pterosid C, c (3S)-pterosid D, d (2R,3R)-pterosin C, e (2R)-pterosid B, f (2S,3R)-pterosin C och g (2R)-pterosin B i den aktiva platsen för BChE. Färgerna på de streckade linjerna förklarar typerna av olika interaktioner: vätebindningsinteraktioner (grönt), hydrofoba interaktioner (rosa) och π-anjoninteraktioner (gyllene). BChE butyrylkolinesteras
Den dockade positionen för 3F9 visade en B.E. på -8,49 kcal/mol. Som visas i figur 8a var hydrofoba interaktioner huvudsakligen ansvariga för ligandbindningen. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329 och Tyr332 stod för de hydrofoba interaktionerna, medan Asp70 visade en π-anjoninteraktion. Figur 8b-e illustrerar de dockade positionerna för (2R,3R)-pterosid C, (3S)-pterosid D, (2R,3R)-pterosin C och (2R)-pterosid B (BChE-hämmare av blandad typ). Deras bindningsficka bestod av Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438 och Tyr440. I enlighet med deras aktivitetsnivåer uppvisade (2R,3R)-pterosid C (IC50 = 5,29 µM), (3S)-pterosid D (IC50 = 19,3 µM), (2R,3R)-pterosin C (IC50 = 20,3 µM) och (2R)-pterosid B (IC50 = 62,0 µM) en B.E. på -7,23, -6,59, -6,52 respektive -4,38 kcal/mol. Som framgår av figur 8b visade 3-OH-gruppen i indanonringen i (2R,3R)-pterosid C två vätebindningar med Gly78 och Tyr440 på avstånden 2,87 respektive 2,89 Å. När det gäller (3S)-pterosid D påverkade 2,2-dimetylgruppen vid indanonringen 3-OH-gruppens bindningsinteraktioner (fig. 8c). 3-OH-gruppen bildade endast en vätebindning med Trp82 på ett avstånd av 2,94 Å, vilket resulterade i en jämförelsevis lägre aktivitet hos (3S)-pterosid D än hos (2R,3R)-pterosid C. Som framgår av fig. 8d misslyckades (2R,3R)-pterosin C med att bilda en vätebindning med Ser287 på grund av avsaknaden av en 2-hydroximetyl-tetrahydropyran-3,4,5-triolgrupp. Följaktligen gav den en lägre aktivitet än (2R,3R)-pterosid C. Avsaknaden av OH-gruppen i position 3 i indanonringen visade sig vara ansvarig för den mycket låga aktiviteten hos (2R)-pterosid B (Fig. 8e), som inte lyckades bilda vätebindningar med Gly78 och Tyr440 på samma sätt som (2R,3R)-pterosid C.
De dockade poserna för (2S,3R)-pterosin C och (2R)-pterosin B (icke-kompetitiva BChE-hämmare) visas i fig. 8f, g respektive. Dessa dockade positioner fanns i det hålrum som omges av Gly283, Leu286, Val288, Phe357 och Asn397. Enligt deras aktivitetsnivåer uppvisade (2S,3R)-pterosin C (IC50 = 44,3 µM) och (2R)-pterosin B (IC50 = 48,1 µM) en B.E. på -5,40 respektive -5,06 kcal/mol. (2S,3 R)-Pterosin C var mer potent än (2R)-pterosin B på grund av närvaron av ytterligare en OH-grupp i position 3 i indanonringen, som bildade en vätebindning med CO-gruppen i Asn397 på ett avstånd av 2,05 Å (fig. 8f). Andra interaktioner visade sig likna (2R)-pterosin B. Som framgår av fig. 8g bildade OH-gruppen i hydroxyetylgruppen i indanonringens position-6 en vätebindning med Gly283 på ett avstånd av 2,40 Å. Resterna Leu286, Val288 och Phe357 deltog i hydrofoba interaktioner.
Hämmare av blandad typ binder till både det fria enzymet och enzymet-substratkomplexet, vilket tyder på att dessa föreningar kan binda till den katalytiska platsen för varje motsvarande enzym. Icke-kompetitiva hämmare binder till den allosteriska platsen i det fria enzymet eller enzym-substratkomplexet. En nyligen genomförd studie tyder på att kompetitiva hämmare, hämmare av blandad typ och icke-kompetitiva hämmare upptar olika platser i bindningsfickorna för BACE1, AChE och BChE38. Under dockning för utvärdering av den hämmande mekanismen hos pterosinderivat definierades föreningarnas bindningsställen i enlighet med deras typ av hämning. Dockningsresultaten visade att bindningsställena för blandade typer och icke-kompetitiva hämmare för BACE1, AChE och BChE delvis överlappar varandra vid varje motsvarande aktiv plats och överensstämde med en tidigare rapport38.
BBB-permeabilitet
PAMPA-BBB, en in vitro artificiell membranpermeabilitetsanalys för BBB, är ett av de mest tillförlitliga fysikalisk-kemiska screeningverktygen i det tidiga skedet för att hitta läkemedel som är inriktade på CNS40. PAMPA-BBB-systemet modellerar den transcellulära passiva spridningen av kemikalier genom BBB och mäter strikt passiva transportmekanismer via ett artificiellt lipidmembran på effektiv permeabilitet (Pe, cm/s). På grundval av det mönster som fastställts för förutsägelse av BBB-permeation klassificerades substanserna i) ”CNS+” (hög förutsedd BBB-permeation); Pe (10-6 cm/s) > 4,00, ii) ”CNS-” (låg förutsedd BBB-permeation); Pe (10-6 cm/s) < 2,00, och iii) ”CNS+/-” (osäker BBB-permeation); Pe (10-6 cm/s) från 4,00 till 2,00. Följaktligen uppvisade (2R)-pterosin B, (2S)-pterosin P och (2S)-pterosin A hög BBB-permeation med Pe-värden på 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s respektive 6,26 × 10-6 cm/s (tabell 4, kompletterande information 4). Pe-värdet för (2R)-pterosin B var 1,7 gånger högre än Pe-värdet för CNSläkemedlet verapamil (Pe = 34,6 × 10-6 cm/s), som användes som positiv kontroll. (2S,3R)-Pterosin C och (2R,3R)-pterosin C uppvisade en osäker BBB-permeation med Pe-värden på 2,34 respektive 1,98. (2R,3R)-Pterosid C, (3S)-Pterosid D och (2R)-Pterosid B, som visade de mest potenta BACE1- och kolinesterashämmande aktiviteterna bland de testade pterosinderivaten, uppvisade en mycket låg BBB-permeabilitet. Förekomsten av 2-hydroximetyl-tetrahydro-pyran-3,4,5-triolgruppen i pterosidderivaten resulterade i en anmärkningsvärd minskning av BBB-permeabiliteten. Jämfört med (2R)-pterosin B minskade den ytterligare närvaron av OH-gruppen i position 3 av indanonringen som i pterosin C, hydroximetylgruppen i position 2 av indanonringen som i (2S)-pterosin A eller hydroximetylgruppen i position 5 av indanonringen som i (2S)-pterosin P också signifikant BBB-permeabiliteten. Med tanke på den exceptionellt höga permeabiliteten i BBB och den betydande hämmningen av BACE1, AChE och BChE kan (2R)-pterosin B ha potential att uppvisa en stark anti-AD-aktivitet.
Effekter av (2R)-pterosin B och (2R,3R)-pterosid C på utsöndringen av Aβ-peptider från neuronala celler
För att undersöka funktionen av (2R)-pterosin B och (2R,3R)-pterosid C när det gäller att minska utsöndringen av Aβ från neuronala celler använde vi en murin neuroblastomcellinje som stabilt överuttrycker humant APPswe. Cellinjen är en cellulär modell av AD som kännetecknas av överdriven utsöndring av Aβ40 och Aβ42. Giftiga amyloidoligomerer bildas av de två isoformerna av Aβ-peptiden med olika längd. Aβ40 är den vanligaste Aβ-isoformen i hjärnan41, medan Aβ42 ökar betydligt med vissa former av AD42. Sandwich ELISA av Aβ40 visade att (2R)-pterosin B signifikant minskade mängden Aβ40-peptid som utsöndrades från neuroblastomcellerna till media med upp till 50 % vid 500 μM (P < 0,01) (fig. 9a). På samma sätt minskade utsöndringen av Aβ42-peptid från neuroblastomcellerna signifikant i närvaro av 500 µM (2R,3R)-pterosin C (P < 0,05) (fig. 9b). Sammanfattningsvis minskade (2R)-pterosin B och (2R,3R)-pterosid C signifikant utsöndringen av Aβ-peptider från neuroblastomceller vid en koncentration på 500 μM.

a Effekten av (2R)-pterosin B på utsöndringen av Aβ-peptider. APPswe-sekreterande neuroblastomceller behandlades med 12, 60, 250 eller 500 µM (2R)-pterosin B i 24 timmar, och konditionerade medier samlades upp i närvaro av proteashämmare. β-SI (10 µM) användes som positiv kontroll. Negativ kontroll omfattade celler som odlats i frånvaro av testföreningar. Kvantitativ analys av utsöndrat Aβ40 och Aβ42 i de konditionerade medierna utfördes med hjälp av sandwich ELISA. Medelvärden ± SD från tre oberoende experiment visas. De utsöndrade Aβ40-peptiderna minskade signifikant i närvaro av 500 µM (2R)-pterosin B. CON negativ kontroll, β-SI β-sekretashämmare III, Aβ β-amyloid. *P < 0,05, **P < 0,01. b Effekt av (2R,3R)-pterosin C på utsöndringen av Aβ-peptider. APPswe-sekreterande neuroblastomceller behandlades med 12, 60, 125, 250 eller 500 µM (2R,3R)-pterosid C i 24 timmar, och konditionerade medier samlades upp i närvaro av proteashämmare. β-SI (10 µM) användes som positiv kontroll. Negativ kontroll omfattade celler som odlats i frånvaro av testföreningar. Kvantitativ analys av utsöndrat Aβ40 och Aβ42 i de konditionerade medierna utfördes med hjälp av sandwich ELISA. Medelvärden ± SD från tre oberoende experiment visas. Den utsöndrade Aβ42-peptiden minskade signifikant i närvaro av 500 µM (2R,3R)-pterosid C. CON negativ kontroll, β-SI β-sekretashämmare III, Aβ β-amyloid. *P < 0.05
Cytotoxicitet hos pterosinderivat baserat på MTT-analys43
Pterosinderivat uppvisade sammantaget försumbar cytotoxicitet mot olika normala och cancercellinjer, t.ex. mänskligt neuroblastom SH-SY5Y, C6-gliaceller från råtta, NIH3T3-fibroblaster från musembryon och B16F10-musmelanom med LD50-värden över 0.5 mM (kompletterande information 5). Särskilt (2R)-pterosin B, (3R)-pterosin D, (2S)-pterosin P, (2S)-pterosid A, (2R)-pterosid B och (2R,3R)-pterosid C uppvisade ingen cytotoxicitet mot de testade cellinjerna med LD50-värden över 5 mM. Intressant nog visade flera pterosiner relativa antiproliferativa effekter mot SH-SY5Y neuronala celler jämfört med C6-gliaceller av mesenkymalt ursprung. Dessa resultat stämmer överens med en tidigare rapport som tyder på att pterosinderivat inte är giftiga för människor26.