
Isolation and characterization of pterosin derivatives from P…. P. aquilinum
P. aquilinumの全草からpterosin誘導体を単離した。 この植物化学成分を調べるために,水抽出物を酢酸エチル (EtOAc) と n-ブタノール (n-BuOH) で順次分配した (Fig. 1). EtOAc可溶分からカラムクロマトグラフィーを繰り返し、(2R)-pterosin B(1)、pterosin Z(2)、(2S)-pterosin P(3)など9つの誘導体が単離された。 (3R)-pterosin D (4), (2S)-pterosin A (5), (2S,3R)-pterosin C (6), (2R,3R)-pterosin C (7), (2R)-pteroside B (8), and pteroside Z (9), with greater than 97% (図)純度。 2). n-BuOH可溶性画分をカラムクロマトグラフィーで繰り返し分離した結果、(2S)-pteroside A (10), (2S)-pteroside A2 (11), (2S,3R)-pteroside C (12), (2R,3R)-pteroside C (13), (3S)-pteroside D (14), (2S)-pteroside P (15) など6種類の派生物を純度97%以上で単離することに成功した。 これらの化合物の構造は、MS、1D-および2D-NMRなどのスペクトルデータの解析により同定された(補足情報1)

プテロシン化合物1〜15xxxの構造
プテロシン誘導体のBACE1、AChEに対する阻害活性を示す。 BChE
抗ADの可能性を評価するために、各プテロシン化合物のBACE1およびコリンエステラーゼに対する阻害活性をそれぞれのin vitro阻害試験で評価した(表1)。 ポジティブコントロールとして用いたケルセチンのIC50が18.8μMであるのに対し、(2S)-pterosin Aと(2S)-pteroside Pは試験した濃度では不活性であったが、すべての試験したプテロシン誘導体はBACE1に対して濃度依存的に阻害作用を示し、IC50値(最大阻害濃度の半分)の範囲は9.74-94.4μMであった。 最も強い阻害剤の阻害効力は、(2R,3R)-pteroside C, (3S)-pteroside D, (2R)-pteroside B, (2S,3R)-pterosin C, (2R,3R)-pterosin C, (2S,3R)-pteroside C, and (2R)-pterosin Bの順でIC50値が9であり、IC50値は2.8μMであった。74, 10.7, 18.0, 23.1, 26.2, 28.9, 29.6 μMであった。 続いて,pterosin誘導体のAChEに対する阻害能を検討した。 陽性対照として用いたberberineのAChEに対するIC50が0.39 μMであるのに対し、試験化合物はいずれも2.55-110 μMの範囲で有意なAChE阻害活性を示した。 AChEに対して最も強い阻害活性を示したプテロシン化合物は、(2R)-pteroside B, (2R,3R)-pteroside C, (2S,3R)-pteroside C, (2S,3R)-pterosin C, および (2R)-pterosin Bで、それぞれIC50値は 2.55, 3.77, 9.17, 12.8, および 16.2 μMであった。 最後に,pterosin誘導体のBChEに対する阻害能について検討した. 試験したすべてのプテロシン化合物はBChEに対して阻害活性を示し、IC50値は5.29から119μMであった。ただし、(3R)-プテロシンDはBChEに対するベルベリンのIC50が3.32μMであるのに対し、試験濃度で不活性であった。 BChEに対して最も強い阻害活性を示したプテロシン化合物は(2R,3R)-pteroside Cとpteroside Zで、IC50値はそれぞれ5.29と5.31μMであった。
まとめてみると、試験したpterosin誘導体のほとんどはBACE1、AChE、BChEに対して同時に大きな阻害活性を示していることが分かった。 プテロシド誘導体のように2-ヒドロキシメチル-テトラヒドロ-ピラン-3,4,5-トリオール基が追加で存在すると、酵素に対する阻害活性が有意に増加した。 さらに、(2R)-pterosin Bのインダノン環の2位に(2S)-pterosin Aと同様にヒドロキシメチル基を付加したものや、(3R)-pterosin Dやpterosin Zのようにメチル基があると酵素に対する阻害活性が減少することが示された。 特に、(2S)-pterosin Pのようにインダノン環の5位にヒドロキシメチル基があるとBACE1に対する阻害活性が低下した。
プテロシン誘導体による酵素阻害の動力学パラメータ
プテロシン誘導体の酵素阻害の様式を説明すべく、BACE1やコリンエステラーゼの代表的阻害剤に対して動力学解析を行った(表2、補足説明2参照)。 Ki(阻害定数)が低いほど、酵素との結合が強固であり、阻害効果が高いことを示す。 全体的に、化合物のKi値はそれぞれのIC50値と相関していた。 化合物 (2R,3R)-pteroside C, (3S)-pteroside D, (2R,3R)-pterosin C による BACE1 抑制は混合型で、Ki 値は 12.6, 16.5, 27.5 であった。6 µMであり、(2R)-pteroside B, (2S,3R)-pterosin C, (2R)-pterosin Bによる阻害は非競合型でKi値はそれぞれ 23.1, 33.8, 38.3 µMであった。 (2R)-pteroside B, (2R,3R)-pteroside C, (2R)-pterosin B, (2S,3R)-pterosin C, (3S)-pteroside DによるAChE阻害は混合型でKiは4.1, 4.0µMであり、typeが異なる。89, 8.13, 12.1, 16.3, 23.1 µMであり、(2R,3R)-pterosin Cは非競合型の阻害剤でKi値は29.6 µMであった。 (2R,3R)-pterosin C, (2R,3R)-pteroside C, (3S)-pteroside D, (2R)-pteroside BによるBChE阻害は混合型でKi値は4.77, 9.62, 19.一方、(2S,3R)-pterosin Cと(2R)-pterosin Bは非競合阻害剤でKi値はそれぞれ29.9と53.5 μMであった。 これらの結果は、特定のプテロシン誘導体が有効なBACE1、AChE、BChE阻害剤となる可能性を示唆するものであった。
BACE1, AChE, BChEの分子ドッキングシミュレーション
BACE1, cholinesterasesはいくつかの結晶構造が利用されています。 野生型構造、共結晶化リガンド、構造の解像度に基づき、ヒトPDBを選択した。 BACE1とQUDの複合体(PDB code: 2WJO, resolution: 2.5 Å)33、AChEとE2020の複合体(PDB code: 4EY7, resolution: 2.35 Å)34、BChEと3F9の複合体(PDB code: 4TPK, resolution: 2.70 Å)35をドッキング用にX線結晶構造として選択しました。 まず、QUD、E2020、3F9を結晶構造から抽出し、それぞれBACE1、AChE、BChEの活性部位にリドッキングした。 その後、BACE1、AChE、BChEに対する既知の阻害機構を持つ(2R,3R)-pteroside C、(3S)-pteroside D、(2R,3R)-pterosin C、(2R)-pterosin Bと、SAR決定のためにドッキングを行った。 ドッキング結果を表 3 にまとめた。 選択した混合型および非競合型のBACE1、AChE、BChE阻害剤のSARにより、プテロシンによる酵素の新規結合部位特異的阻害を評価することが可能となった。
E2020のドッキングモードは、リコンビナントのヒトAChE(rhAChE)34について報告済みの実験的に決定した結合モードと一致していました。 E2020の結晶とドッキングしたコンフォメーションの二乗平均平方根偏差(RMSD)は0.54Åであり、実験的結合様式を再現する我々のドッキングセットアップの信頼性が示唆されました。 また、E2020のドッキングモードは、rhAChE-E2020と同様の相互作用につながりました。 本研究では、ドッキングの際に結晶構造から水分子を取り除いたため、水を介した相互作用は解析していない。 同様に、QUDと3F9のドッキングモードは、それぞれBACE1 33とBChE35の利用可能な実験データと一致した(補足情報3)。 QUDと3F9の結晶とドッキングしたコンフォメーション間のRMSDは、それぞれ0.46と0.60Åであった。 さらに、プテロシン阻害剤の結合部位は、BACE1、AChE、BChE38を対象とした過去のドッキング研究とも一致した。 ただし、この研究ではTetronarce californica AChE (PDB code: 1ACJ) を使用しており、ヒトAChEとは配列が異なるため残基番号が若干異なっている。
BACE1 ドッキング
阻害タイプと活性に基づいて、混合型および非競合型BACE1阻害剤のドッキングモードを示すため、それぞれ (2R,3R) -pteroside Cと (2R) -pteroside Bが選択されました。 図 3a, b は、それぞれ (2R,3R)-pteroside C と (2R)-pteroside B のドッキングモデルである。 図3:混合型および非競合型BACE1阻害剤の分子ドッキングモデル。

aは混合型BACE1阻害剤(2R、3R)-pteroside C(青色)、bは非競合型BACE1阻害剤(2R)-pteroside B(茶色)に対する分子ドッキングモデルである。 ドッキングポーズは、QUDのX線結晶構造(黄色)と重ね合わせた(PDBコード:2WJO)。 BACE1、活性部位残基、化合物はそれぞれリボン、ライン、スティックモデルで示した。 点線の色は、水素結合相互作用(緑)、疎水性相互作用(ピンク)、πシグマ相互作用(紫)という様々な相互作用の種類を表している。 BACE1 βサイトアミロイド前駆体タンパク質切断酵素1

BACE1の活性部位におけるa QUD、b (2R,3R)-pteroside C、c (3S)-pteroside D、d (2R,3R)-pterosin C、e (2R)-pteroside B、f (2S,3R)-pterosin Cおよびg (2R)-pterosin Bのリガンドインタラクション図。 点線の色は、水素結合相互作用(緑)、疎水性相互作用(ピンク)、πシグマ相互作用(紫)と、様々な相互作用の種類を表している。 BACE1 βサイトアミロイド前駆体タンパク質切断酵素1
QUDのドッキングポーズは-7.59 kcal/mol の結合エネルギー(B.E.)を示していました。 図4aに示すように、リガンドのキナゾリン環上のNH2基はAsp32とAsp228のCO基とそれぞれ1.86と2.17Åの距離で2つの水素結合を見せた。 Leu30、Tyr71、Phe108、Val332は疎水性相互作用の仲介役であった。 図 4b-d は、混合型 BACE1 阻害剤である (2R,3R)-pteroside C、 (3S)-pteroside D および (2R,3R)-pterosin C のドッキングポーズをそれぞれ示したものであ る。 これらはSer36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228, Thr231で囲まれた結合ポケットに配置されていることが確認された。 活性値としては、(2R,3R)-pteroside C (IC50 = 9.74 µM), (3S)-pteroside D (IC50 = 10.7 µM), (2R,3R)-pterosin C (IC50 = 26.2 µM) がそれぞれ -7.27, -6.93 および -4.84 kcal/modalとなり、BASEは2.0であることが示された。 (2R,3R)-Pteroside Cは(2R,3R)-pterosin Cよりも高い効力を示したが、これは3つの水素結合を形成する2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol基が追加で存在しているためである (Fig. 4b). ヒドロキシメチルのOH基とテトラヒドロピラントリオール環の3-OH基は、Asp228のCO基とそれぞれ2.63と2.12 Åの距離で2つの水素結合を形成した。 3S)-プテロシドD(図4c)では、インダノン環の2,2-ジメチル基の存在が(2R,3R)-プテロシドCと比較して結合相互作用をわずかに変化させており、テトラヒドロピラントリオール環の3-OHグループはAsp228と1.93 Åの距離に1つの水素結合のみを示している。 しかし,インダノン環の3-OH基はIle126と2.34Åの距離で水素結合を示した。これらの相互作用は(2R,3R)-pteroside Cと比較して(3S)-pteroside Dの活性をわずかに減少させた。 図 4d に示した (2R,3R)-pterosin C のドッキング相互作用では、インダノン環の 3-OH 基が Ser36 と Asn37 にそれぞれ 1.83 と 2.07Å の距離で水素結合を介し て結合していることがわかった。 Ala39、Val69、Trp76、Ile118、Arg128は疎水性相互作用を示し、Val69はπシグマ相互作用を示した。
(2R)-pteroside B, (2S,3R)-pterosin C, (2R)-pterosin B (noncompetitive BACE1 inhibitor) をそれぞれ図 4e-g にドッキングしたポーズを示した。 これらは、Asn37, Val69, Tyr71, Trp76, Lys107, Phe108, Ile126で囲まれたキャビティにドッキングしていることが確認された。 活性値から、(2R)-pteroside B (IC50 = 18.0 µM), (2S,3R)-pterosin C (IC50 = 23.1 µM), および (2R)-pterosin B (IC50 = 29.6 µM) はそれぞれ -6.16, -5.07, および -4.64 kcal/mol という B.E. を示していることがわかった。 Fig. 4e に示すように、(2R)-pteroside B は (2S,3R)-pterosin C および (2R)-pterosin B より高い活性を示したが、これは 4 つの水素結合相互作用を示した追加の 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol 基の存在に起因している。 テトラヒドロピラントリオール環の4-OH基とAsn37のNH基およびCO基の間には、それぞれ2.48と2.19Åの距離で2つの水素結合が観測された。 さらに、3-OH基と5-OH基はIle126とTrp76とそれぞれ2.19Åと2.46Åの距離で追加の水素結合を見せた。 (2S,3R)-Pterosin Cは(2R)-pterosin Bよりもわずかに優れた活性を示したが、これはインダノン環の3位に追加のOH基が存在し、Lys107と2.12Åの距離で水素結合を形成しているためである(Fig. 4f)。 その他の相互作用は(2R)-pterosin Bと同様であった。図4gに示すように、(2R)-pterosin BはVal69, Tyr76, Phe108と疎水的相互作用を示していた。
AChE ドッキング
(2R)-Pteroside B と (2R,3R)-pterosin C は、その活性と AChE 阻害の種類から、それぞれ混合型と非競合型 AChE 阻害剤のドッキング様式を示す代表として選択された。 図 5a, b は、それぞれ (2R)-pteroside B と (2R,3R)-pterosin C のドッキングモデ ルを示すものである。 図 5:混合型および非競合型 AChE 阻害剤の分子ドッキングモデル。

a 混合型AChE阻害剤(2R)-pteroside B(青色)とb 非競合型AChE阻害剤(2R,3R)-pterosin C(茶色)に関する分子ドッキングモデル. ドッキングしたポーズはE2020のX線結晶構造(黄色)と重ね合わせた(PDBコード:4EY7)。 AChE、活性部位残基、化合物はそれぞれリボン、ライン、スティックモデルで示されている。 点線の色は、水素結合相互作用(緑)、疎水性相互作用(ピンク)という様々な相互作用の種類を説明している。 AChEアセチルコリンエステラーゼ

AChEの活性部位におけるa E2020、b (2R)-pteroside B、c (2R,3R)-pteroside C、d (2S,3R)-pterosin C、e (2R)-pterosin B、f (3S)-pteroside D、g (2R,3R)-pterosin Cのリガンドインタフェースダイアグラムである。 点線の色は、水素結合相互作用(緑)、疎水性相互作用(ピンク)、πシグマ相互作用(紫)と、様々な相互作用の種類を表している。 AChE acetylcholinesterase
E2020のドッキングポーズは-10.28 kcal/molのB.E.を示しました。 図6aに示すように、インダノン環のCO基はPhe295のNH基と1.70Åの距離で水素結合を形成し、Trp286とTyr341はπ-シグマ相互作用に関与し、Trp86, Trp286, Tyr337, Phe338, および Tyr341は疎水性相互作用を媒介した。 図 6b-f は、(2R)-pteroside B, (2R,3R)-pteroside C, (2S,3R)-pterosin C, (2R)-pterosin B および (3S)-pteroside D(混合型 AChE 阻害剤)のドッキング姿勢をそれぞれ示したものであ る。 これらはTyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338, Tyr341で囲まれた活性部位に収容された。 その活性値と一致して、(2R)-pteroside B (IC50 = 2.55 µM), (2R,3R)-pteroside C (IC50 = 3.77 µM), (2S,3R)-pterosin C (IC50 = 12. 12).8 µM)、(2R)-pterosin B(IC50 = 16.2 µM)および(3S)-pteroside D(IC50 = 27.4 µM)はそれぞれ -7.90, -7.49, -6.03, -5.76 および -4.91 kcal/mol という B.E. を示 した。 (2R)-プテロシドBが(2S,3R)-プテロシンCおよび(2R)-プテロシンBより高い効力を示したのは,さらに2-ヒドロキシメチル-テトラヒドロ-ピラン-3,4,5-トリオール基が存在して,3つの水素結合相互作用が確立しているためである(図6b)。 ヒドロキシメチルのOH基とテトラヒドロピラントリオール環の3-OH基は、Asn87のCO基とそれぞれ2.23と2.28 Åの距離で水素結合を形成した。 (2R,3R)-pteroside C (Fig. 6c) の場合、インダノン環の2位のメチル基はTyr341と疎水的相互作用を示さず、(2R)-pteroside Bよりも比較的低い活性を示している。 (2S,3R)-pterosinCが(2R)-pterosinBより活性が高いのは、インダノン環の3位にOH基が存在し、Phe295と1.82 Åの距離で水素結合を形成しているためである(図6d)。 残りの相互作用は、(2R)-pterosin Bの相互作用と同等であった。 図6eに示すように、インダノン環の6位にあるヒドロキシエチル基のOH基はTyr124と2.48Åの距離で水素結合を形成した。Tyr124, Trp286, Tyr337, Phe338, Tyr341は疎水性相互作用に寄与していた。 (2R)-プテロシドBや(2R,3R)-プテロシドCと比較して、(3S)-プテロシドDはインダノン環に2,2-ジメチル基があるため異種の結合相互作用を示した(図6f)。 テトラヒドロピラントリオール環の5-OH基はAsp74と1.96 Åの距離で水素結合を形成していた。 これらの相互作用が(3S)-pteroside Dの低い活性を説明した。
(2R,3R)-pterosin C(非競合的AChE阻害剤)のドッキング姿勢を図6gに表示した。 (2R,3R)-Pterosin C (IC50 = 23.2 µM) の B.E. は -5.01 kcal/mol であることが示された。 (2R,3R)-pterosin Cの結合ポケットはTyr72, Trp286, Ser293, Phe297, Tyr341からなり、2つの水素結合相互作用を持っていた。 一つはインダノン環の3-OH基とSer293のCO基の間に2.03Åの距離で形成され、もう一つはインダノン環の6位に存在するヒドロキシエチル基のOH基とTyr72の間に1.94Åの距離で水素結合が観察されている。 また、Tyr72, Trp286, Phe297, Tyr341などの残基が疎水性相互作用に関与していた。
BChE ドッキング
BChE阻害の活性レベルと種類を考慮し、混合型および非競合型BChE阻害剤のドッキング様式を示すため, それぞれ、 (2R,3R)-pteroside Cと (2S,3R)-pterosin Cが選択されていた。 図 7a, b は、それぞれ (2R,3R)-pteroside C と (2S,3R)-pterosin C のドッキングモデ ルを示したものである。 図 7:混合型および非競合型 BChE 阻害剤の分子ドッキングモデル。
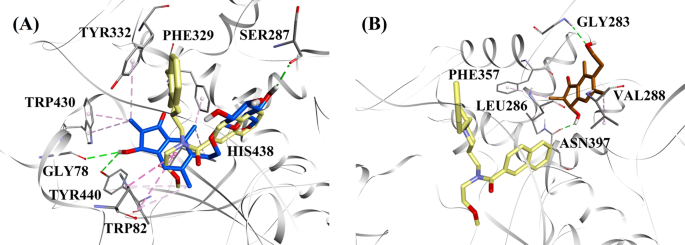
aは混合型BChE阻害剤(2R,3R)-pteroside C(青色)、bは非競合型BChE阻害剤(2S,3R)-pterosin C(茶色)に対する分子ドッキングモデルである。 ドッキングしたポーズは3F9のX線結晶構造(黄色)(PDBコード:4TPK)と重ね合わせた。 BChE、活性部位残基、化合物はそれぞれリボン、ライン、スティックモデルで示した。 点線の色は、水素結合相互作用(緑色)、疎水性相互作用(ピンク色)という様々な相互作用の種類を説明している。 BChEブチリルコリンエステラーゼ
BChEの活性部位におけるa 3F9, b (2R,3R)-pteroside C, c (3S)-pteroside D, d (2R,3R)-pterosin C, e (2R)-pteroside B, f (2S,3R)-pterosin C and g (2R)-pterosin Bのリガンド相互作用図である。 点線の色は、水素結合相互作用(緑)、疎水性相互作用(ピンク)、π-アニオン相互作用(金色)と、様々な相互作用の種類を表している。 BChE butyrylcholinesterase
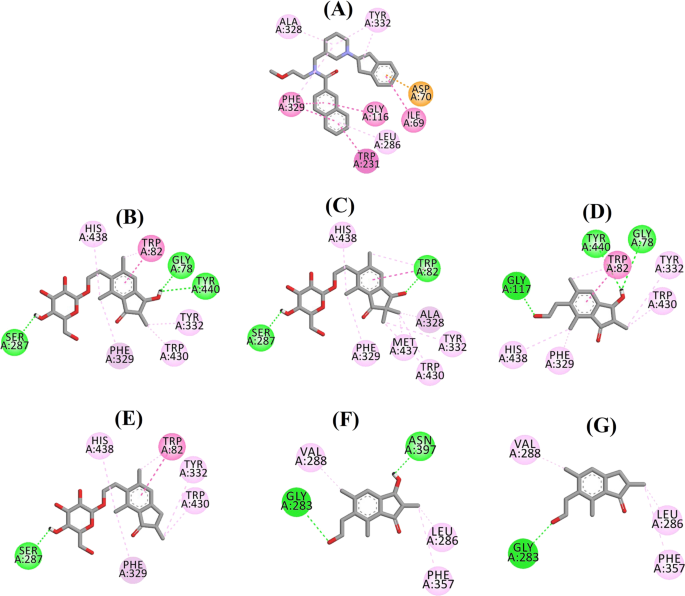
3F9のドッキングポーズは、-8.49 kcal/molのB.E.を示しました。 図8aに示すように、リガンド結合には主に疎水性相互作用が関与していた。 Ile69, Gly116, Trp231, Leu286, Ala328, Phe329, Tyr332 が疎水性相互作用を、Asp70 がπ-アニオン相互作用を担っていた。 図 8b-e は、混合型 BChE 阻害剤である (2R,3R)-pteroside C、 (3S)-pteroside D、 (2R,3R)-pterosin C および (2R)-pteroside B のドッキングポーズをそれぞれ示したものである。 これらの結合ポケットはGly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438, Tyr440で構成されていた。 その活性レベルに応じて、(2R,3R)-pteroside C (IC50 = 5.29 µM), (3S)-pteroside D (IC50 = 19.3 µM), (2R,3R)-pterosin C (IC50 = 20.3 µM) および (2R)-pteroside B (IC50 = 62.0 µM) はそれぞれ -7.23, -6.59, -6.52 および -4.38 kcal/mol という B.E. を実証している。 Fig. 8b に示すように、(2R,3R)-pteroside C のインダノン環の 3-OH 基は、Gly78 および Tyr440 とそれぞれ 2.87 および 2.89Å の距離で水素結合を形成していることがわかった。 (3S)-pterosideDの場合、インダノン環の2,2-ジメチル基が3-OH基の結合相互作用に影響を与えた(図8c)。 図8dに示すように、(2R,3R)-pterosin Cは2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol基がないため、Ser287と水素結合を作ることができない。 また、(2R)-pteroside B の活性が非常に低いのは、インダノン環の 3 位に OH 基がないためであることが判明した(Fig. 8e)は、(2R,3R)-pteroside Cと同様にGly78およびTyr440と水素結合を形成できなかった。
(2S,3R)-pterosin Cおよび(2R)-pterosin B(非競合的BChE阻害剤)のドッキングポーズをそれぞれ図8f、gに示す。 これらのドッキングポーズは、Gly283, Leu286, Val288, Phe357, Asn397 で囲まれたキャビティに収まっていた。 活性値から、(2S,3R)-pterosin C (IC50 = 44.3 µM) と (2R)-pterosin B (IC50 = 48.1 µM) はそれぞれ -5.40 と -5.06 kcal/mol という B.E. を示している。 (2S,3 R)-プテロシン C は (2R)-pterosin B よりも強力だったが、これはインダノン環の 3 位にさらに OH 基があり、Asn397 の CO 基と 2.05 Å の距離で水素結合を形成していたからである(図 8f)。 その他の相互作用は(2R)-pterosin Bと同様であった。Fig. 8gに示すように、インダノン環の6位にあるヒドロキシエチル基のOH基は、Gly283と2.40 Åの距離で水素結合を形成していることが確認された。 また,Leu286, Val288, Phe357は疎水的な相互作用を形成していた。
混合型阻害剤は遊離酵素と酵素-基質複合体の両方に結合することから,それぞれの酵素の触媒部位に結合することが示唆される。 非競合型阻害剤は遊離酵素あるいは酵素-基質複合体のアロステリック部位に結合する。 最近の研究では、競合型、混合型、非競合型の阻害剤は、BACE1、AChE、BChE38の結合ポケットにおいて異なる部位を占めることが示唆された。 pterosin 誘導体の阻害機構を評価するためのドッキングの際に、阻害のタイプに応じて化合物の結合部位を定義した。 その結果、BACE1、AChE、BChEに対する混合型阻害剤および非競合型阻害剤の結合部位は、対応する各活性部位で互いに一部重複しており、以前の報告38と一致した。
BB permeability
PAMPA-BBB, in vitro人工膜透過性アッセイは中枢神経標的薬早期探索において最も信頼できる物理化学スクリーニングツール40の1つである。 PAMPA-BBBシステムは、BBBを介した化学物質の細胞内受動拡散をモデル化し、人工脂質膜を介した受動輸送メカニズムを有効透過率(Pe、cm/s)で厳密に測定する。 BBB透過予測のために確立されたパターンに基づいて、化合物は、(i) “CNS+” (高いBBB透過予測); Pe (10-6 cm/s) > 4.00, (ii) “CNS-” (低いBBB透過予測); Pe (10-6 cm/s) < 2.00, (iii) “CNS+/-“(BBB透過不確か); Pe (10-6 cm/s) 4.00 to 2.00 のように分類された。 その結果、(2R)-pterosin B、(2S)-pterosin P、(2S)-pterosin AはそれぞれPe値が60.3 × 10-6 cm/s, 7.92 × 10-6 cm/s, 6.26 × 10-6 cm/sで高いBBB透過性を示した(表4、補足説明4)。 2R)-プテロシンBのPe値は、陽性対照に用いた中枢神経系薬剤であるベラパミル(Pe=34.6×10-6cm/s)のPe値の1.7倍であった。 (2S,3R)-Pterosin Cおよび(2R,3R)-pterosin CはPeがそれぞれ2.34および1.98であり,不確実なBBB透過を示した。 (2R,3R)-Pteroside C,(3S)-pteroside D,(2R)-pteroside Bは,試験したpterosin誘導体の中で最も強力なBACE1-およびコリンエステラーゼ阻害活性を示したが,非常に低いBBB透過性を示した。 pteroside誘導体のように2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol基が存在することにより、BBB透過性が著しく低下することがわかった。 (2R)-プテロシンBと比較して、プテロシンCのようにインダノン環の3位にOH基、(2S)-プテロシンAのようにインダノン環の2位にヒドロキシメチル基、(2S)-プテロシンPのようにインダノン環の5位にヒドロキシメチル基を追加して含有させた場合も、BBB透過性が大きく低下することが確認された。 BBB透過性が極めて高く、BACE1、AChE、BChEを有意に阻害することから、(2R)-pterosin Bは強い抗AD活性を示す可能性があると考えられる。
(2R)-pterosin Bおよび(2R,3R)-pteroside Cの神経細胞によるAβペプチド分泌への影響
(2R)-pterosin Bおよび (2R.) の機能については、Pterosin誘導体を用いて検討することにしました。3R)-プテロシドCが神経細胞からのAβの排泄を減少させることを、ヒトAPPsweを安定的に過剰発現しているマウス神経芽腫細胞株を用いて明らかにした。 この細胞株は、Aβ40とAβ42の過剰分泌を特徴とするADの細胞モデルである。 有害なアミロイドオリゴマーは、長さの異なる2種類のAβペプチドから形成される。 Aβ40は脳内に最も多く存在するAβ40アイソフォームですが41、Aβ42は特定のADの形態で著しく増加します42。 Aβ40のサンドイッチELISAは、(2R)-pterosin Bが500μMで神経芽腫細胞から培地に分泌されるAβ40ペプチドの量を50%まで著しく減少させた(P < 0.01) (Fig. 9a). 同様に,神経芽細胞腫細胞によるAβ42ペプチドの分泌は,500μMの(2R,3R)-プテロシドCの存在下で有意に減少した(P < 0.05)(図9b)。 以上より、(2R)-pterosin Bおよび(2R,3R)-pteroside Cは、500μMの濃度で神経芽細胞腫細胞からのAβペプチドの分泌を有意に減少させた。

a (2R)-プテロシンBのAβペプチド分泌への影響。 APPsweを分泌する神経芽細胞は、12、60、250、または500μMの(2R)-pterosin Bで24時間処理し、プロテアーゼ阻害剤存在下でコンディショニングメディアを収集した。 β-SI (10 µM)をポジティブコントロールとして使用した。 陰性コントロールは、試験化合物非存在下で培養した細胞を含む。 調整培地中の分泌されたAβ40およびAβ42の定量分析は、サンドイッチELISAを使用して行った。 3つの独立した実験からの平均値±SDを示す。 分泌された Aβ40 ペプチドは、500μM (2R)-pterosin B の存在下で有意に減少した。CON 負コントロール、β-SI β-secretase inhibitor III、Aββ-amyloid.は、分泌された Aβ40 ペプチドは、500μM (2R)-pterosin B の存在下で有意に減少した。 *(2R,3R)-pteroside CがAβペプチドの分泌に及ぼす影響。 APPswe分泌神経芽細胞は、12、60、125、250、または500μMの(2R,3R)-pteroside Cで24時間処理し、プロテアーゼ阻害剤存在下で調整培地を回収した。 β-SI (10 µM)をポジティブコントロールとして使用した。 陰性コントロールには、試験化合物非存在下で培養した細胞を用いた。 サンドイッチELISA法により、調整培地中の分泌型Aβ40およびAβ42を定量的に解析した。 3回の独立した実験からの平均値±SDを示す。 分泌されたAβ42ペプチドは、500μM (2R,3R)-プテロシドCの存在下で有意に減少した。CONネガティブコントロール、β-SI β-セクレターゼ阻害剤 III、Aβ β-アミロイド。 *P < 0.05
MTT assayに基づくプテロシン誘導体の細胞毒性43
全体として、プテロシン誘導体は、SH-SY5Yヒト神経芽腫、C6ラットグリア細胞、NIH3T3マウス胚線維芽細胞およびB16F10マウス黒色腫など様々な正常および癌細胞株に対して無視できる細胞毒性を示し、0以上のLD50値で、その細胞毒性は0.5 mMであった(補足情報5)。 特に、(2R)-pterosin B, (3R)-pterosin D, (2S)-pterosin P, (2S)-pteroside A, (2R)-pteroside B, および (2R,3R)-pteroside CはLD50が5mM以上で細胞毒性を示さなかった。 興味深いことに、いくつかのプテロシンはSH-SY5Y神経細胞に対して、間葉系グリア細胞C6と比較して相対的に抗増殖作用を示した。 この結果は,pterosin誘導体がヒトに対して無毒であることを示唆した過去の報告26.
と一致するものであった.