
Isolamento e caracterização de derivados de pterosina de P. aquilinum
Derivados de pterosina foram isolados das plantas inteiras de P. aquilinum. Para a investigação dos constituintes fitoquímicos da samambaia, um extrato de água foi dividido sucessivamente com acetato de etilo (EtOAc) e n-butanol (n-BuOH) (Fig. 1). A cromatografia em coluna repetida da fração EtOAc-solúvel resultou no isolamento de nove derivados, incluindo (2R)-pterosina B (1), pterosina Z (2), (2S)-pterosina P (3), (3R)-pterosina D (4), (2S)-pterosina A (5), (2S,3R)-pterosina C (6), (2R,3R)-pterosina C (7), (2R)-pterosídeo B (8), e pterosídeo Z (9), com purezas superiores a 97% (Fig. 2). A separação cromatográfica em coluna repetida da fração n-BuOH-solúvel resultou no isolamento de seis derivados, incluindo (2S)-pterosídeo A (10), (2S)-pterosídeo A2 (11), (2S,3R)-pterosídeo C (12), (2R,3R)-pterosídeo C (13), (3S)-pterosídeo D (14) e (2S)-pterosídeo P (15), com purezas superiores a 97%. As estruturas dos compostos foram identificadas pela análise dos dados espectrais, incluindo EM, 1D- e 2D-NMR (Informações Suplementares 1).

Estruturas dos compostos de pterosina 1-15xxx
Atividade inibitória dos derivados de pterosina contra BACE1, AChE, e BChE
Para avaliar o potencial anti-AD, a atividade inibitória de cada composto de pterosina contra BACE1 e colinesterases foi avaliada pelos respectivos ensaios de inibição in vitro (Tabela 1). Todos os derivados de pterosina testados mostraram atividade inibitória dependente da concentração contra BACE1 com uma faixa de valores de IC50 (concentração inibitória semimáxima) de 9,74-94,4 μM, com exceção da (2S)-pterosina A e (2S)-pterosida P que estavam inativos nas concentrações testadas, em comparação com a IC50 da quercetina utilizada como controle positivo, que foi de 18,8 μM. A potência inibitória dos inibidores mais fortes foi da ordem de (2R,3R)-pterosídeo C, (3S)-pterosídeo D, (2R)-pterosídeo B, (2S,3R)-pterosídeo C, (2R,3R)-pterosídeo C, (2S,3R)-pterosídeo C, e (2R)-pterosídeo B com valores de IC50 de 9.74, 10,7, 18,0, 23,1, 26,2, 28,9, e 29,6 μM, respectivamente. Posteriormente testamos os potenciais inibitórios dos derivados da pterosina contra a AChE. Todos os compostos testados mostraram atividades inibitórias de AChE significativas, com valores de IC50 na faixa de 2,55-110 μM, em comparação com a IC50 contra AChE da berberina utilizada como controle positivo, que foi de 0,39 μM. Os compostos de pterosina que apresentaram maior atividade inibitória contra AChE foram (2R)-pterosida B, (2R,3R)-pterosida C, (2S,3R)-pterosida C, (2S,3R)-pterosina C, e (2R)-pterosina B com valores de IC50 de 2,55, 3,77, 9,17, 12,8, e 16,2 μM, respectivamente. Finalmente, nós testamos as capacidades inibitórias dos derivados de pterosina contra BChE. Todos os compostos de pterosina testados mostraram atividade inibitória contra BChE, com valores de IC50 que variaram de 5,29 a 119 μM, com exceção da (3R)-pterosina D que estava inativa nas concentrações testadas, em comparação com a IC50 de berberina contra BChE, que foi de 3,32 μM. Os compostos de pterosina que apresentaram a mais forte atividade inibitória contra BChE foram (2R,3R)-pterosídeo C e pterosídeo Z com valores de IC50 de 5,29 e 5,31 μM, respectivamente.
Coletivamente, a maioria dos derivados de pterosina testados exibiram atividades inibitórias significativas contra BACE1, AChE e BChE simultaneamente. A presença do grupo adicional 2-hidroximetil-tetrahidro-pirano-3,4,5-triol como nos derivados de pterósido aumentou significativamente as atividades inibitórias contra as enzimas. Além disso, a presença do grupo hidroximetil adicional na posição 2 do anel de indanona de (2R)-pterosina B como em (2S)-pterosina A ou do grupo metilo como em (3R)-pterosina D e pterosina Z diminuiu as atividades inibitórias contra as enzimas. Em particular, a presença do grupo hidroximetil na posição 5 do anel indanone como em (2S)-pterosina P diminuiu a atividade inibitória contra BACE1.
Parâmetros cinéticos de inibição enzimática por derivados de pterosina
Na tentativa de explicar o modo de inibição enzimática dos derivados de pterosina, realizamos uma análise cinética de BACE1 e colinesterases para inibidores representativos (Tabela 2, Informações Suplementares 2). Um baixo Ki (constante de inibição) indica uma ligação enzimática mais apertada e um inibidor mais eficaz. Em geral, os valores de Ki dos compostos estão correlacionados com os respectivos valores de IC50. A inibição BACE1 pelos compostos (2R,3R)-pterosídeo C, (3S)-pterosídeo D, e (2R,3R)-pterosídeo C foi do tipo misto com valores de Ki de 12,6, 16,5, e 27.6 µM, respectivamente, enquanto a inibição por (2R)-pterosídeo B, (2S,3R)-pterosídeo C, e (2R)-pterosídeo B foi não competitiva com valores de Ki de 23,1, 33,8, e 38,3 µM, respectivamente. A inibição de AChE por (2R)-pterosida B, (2R,3R)-pterosida C, (2R)-pterosina B, (2S,3R)-pterosina C, e (3S)-pterosida D foi do tipo misto com valores de Ki de 4.89, 8,13, 12,1, 16,3, e 23,1 µM, respectivamente, enquanto (2R,3R)-pterosina C foi um inibidor do tipo não competitivo com um valor de Ki de 29,6 µM. A inibição de BChE por (2R,3R)-pterosina C, (2R,3R)-pterosídeo C, (3S)-pterosídeo D, e (2R)-pterosídeo B foi do tipo misto com valores de Ki de 4,77, 9,62, 19.7, e 22,6 µM, respectivamente, enquanto (2S,3R)-pterosina C e (2R)-pterosina B foram inibidores não competitivos com valores de Ki de 29,9 e 53,5 µM, respectivamente. Assim, estes resultados sugerem que derivados específicos de pterosina podem ser efetivos BACE1, AChE, e inibidores de BChE.
Simulações de acoplamento molecular para BACE1, AChE e BChE
Estruturas de cristais siderais estão disponíveis para BACE1 e colinesterases. Selecionamos PDBs humanos baseados em estruturas do tipo selvagem, ligandos cocristalinos e resoluções das estruturas. Estruturas de cristal de raios X de BACE1 complexadas com QUD (código PDB: 2WJO, resolução: 2.5 Å)33, AChE complexadas com E2020 (código PDB: 4EY7, resolução: 2.35 Å)34, e BChE complexadas com 3F9 (código PDB: 4TPK, resolução: 2.70 Å)35 foram selecionadas para acoplamento. Inicialmente, QUD, E2020 e 3F9 foram extraídos das estruturas de cristal e refazidos nos locais ativos do BACE1, AChE e BChE, respectivamente. Subsequentemente, (2R,3R)-pterosídeo C, (3S)-pterosídeo D, (2R,3R)-pterosídeo C, (2R)-pterosídeo B, (2S,3R)-pterosídeo C e (2R)-pterosídeo B com o conhecido mecanismo de inibição contra BACE1, AChE e BChE foram ancorados para determinar a sua SAR. Os resultados da atracagem estão resumidos na Tabela 3. A SAR dos inibidores de BACE1, AChE e BChE de tipo misto e não competitivo selecionados permitiu a avaliação da nova inibição das enzimas por pterosinas.
Our docking mode of E202020 was consistent with the experimentally determined binding mode previously reported with recombinant human AChE (rhAChE) (Supplementary Information 3)34. O desvio raiz-medo quadrado (RMSD) entre o cristal e as conformações acopladas do E2020 foi de 0,54 Å, o que sugeriu a confiabilidade da nossa configuração de acoplamento na reprodução do modo de ligação experimental. Além disso, o modo acoplado do E2020 levou a uma interação semelhante à do rhAChE-E202020. Em nosso estudo, as moléculas de água foram removidas da estrutura cristalina durante a docagem; portanto, as interações mediadas pela água não foram analisadas no presente estudo. Da mesma forma, os modos ancorados de QUD e 3F9 foram consistentes com os dados experimentais disponíveis para BACE1 33 e BChE35, respectivamente (Informação Suplementar 3). Os RMSDs entre o cristal e as conformações acopladas do QUD e 3F9 foram de 0,46 e 0,60 Å, respectivamente. Além disso, os locais de ligação dos inibidores de pterosina estavam de acordo com um estudo de acoplamento anterior que envolveu BACE1, AChE, e BChE38. Entretanto, o estudo utilizou Tetronarce californica AChE (código PDB: 1ACJ), que contém números de resíduos ligeiramente diferentes do AChE humano devido a variações em suas seqüências.
BACE1 docking
Baseado no tipo de inibição e atividade, (2R,3R)-pterosídeo C e (2R)-pterosídeo B foram selecionados para demonstrar os modos acoplados de inibidores BACE1 mistos e não-competitivos, respectivamente. A Figura 3a, b mostra os modelos acoplados de (2R,3R)-pterosídeo C e (2R)-pterosídeo B, respectivamente. As interações dos compostos acoplados dentro do sítio ativo do BACE1 são mostradas na Fig. 4.

Modelos de acoplamento molecular para o inibidor misto tipo BACE1 (2R,3R)-pterosídeo C (cor azul) e b o inibidor não competitivo tipo BACE1 (2R)-pterosídeo B (cor marrom). Posições bloqueadas são sobrepostas à estrutura de cristal de raios X de QUD (cor amarela) (código PDB: 2WJO). BACE1, resíduos e compostos ativos do local são mostrados pelos modelos de fita, linha e bastão, respectivamente. As cores das linhas pontilhadas explicam os tipos de várias interacções: interacções de ligação de hidrogénio (verde), interacções hidrofóbicas (rosa) e interacções π-sigma (púrpura). BACE1 β-site amilóide precursor da enzima de clivagem da proteína 1

Diagrama de interação ligand de um QUD, b (2R,3R)-pterosídeo C, c (3S)-pterosídeo D, d (2R,3R)-pterosídeo C, e (2R)-pterosídeo B, f (2S,3R)-pterosídeo C, e g (2R)-pterosídeo B no sítio ativo do BACE1. As cores das linhas pontilhadas explicam os tipos de várias interações: interações de ligação de hidrogênio (verde), interações hidrofóbicas (rosa) e interações π-sigma (púrpura). BACE1 β-site amilóide precursor da proteína de clivagem enzima 1
A posição acoplada de QUD exibiu uma energia de ligação (B.E.) de -7,59 kcal/mol. Como mostrado na Fig. 4a, o grupo NH2 no anel de quinazolina do ligante mostrou duas ligações de hidrogênio com os grupos CO dos Asp32 e Asp228 a distâncias de 1,86 e 2,17 Å, respectivamente. Uma terceira ligação de hidrogênio foi observada entre o outro grupo NH do ligante e o grupo CO de Gly230 a uma distância de 2,16 Å. Leu30, Tyr71, Phe108 e Val332 mediaram as interações hidrofóbicas. A Figura 4b-d mostra as poses acopladas de (2R,3R)-pterosídeo C, (3S)-pterosídeo D e (2R,3R)-pterosídeo C (inibidores de BACE1 do tipo misto), respectivamente. Eles foram posicionados na bolsa de ligação forrada por Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228, e Thr231. Conforme seus níveis de atividade, (2R,3R)-pterosídeo C (IC50 = 9,74 µM), (3S)-pterosídeo D (IC50 = 10,7 µM) e (2R,3R)-pterosídeo C (IC50 = 26,2 µM) exibiram um B.E. de -7,27, -6,93, e -4,84 kcal/mol, respectivamente. (2R,3R)-Pterósido C exibiu uma potência maior que (2R,3R)-pterosina C devido à existência de um grupo adicional de 2-hidroximetil-tetra-hidro-pirano-3,4,5-triol, que formou três ligações de hidrogênio (Fig. 4b). O grupo OH do hidroximetil e o grupo 3-OH do anel tetrahidro-piranotriol apresentaram duas ligações de hidrogênio com o grupo CO do Asp228 a distâncias de 2,63 e 2,12 Å, respectivamente. Além disso, o grupo 3-OH demonstrou uma ligação de hidrogênio com Thr231 a uma distância de 1,89 Å. No caso do grupo (3S)-pterosídeo D (Fig. 4c), a presença do grupo 2,2-dimetil no anel de indanona alterou ligeiramente as interações de ligação em relação ao grupo (2R,3R)-pterosídeo C. O grupo 3-OH do anel tetrahidro-piranotriol mostrou apenas uma única ligação de hidrogênio com Asp228 a uma distância de 1,93 Å. Entretanto, o grupo 3-OH do anel indanone exibiu uma ligação adicional de hidrogênio com Ile126 a uma distância de 2,34 Å. Essas interações diminuíram ligeiramente a atividade do (3S)-pterosídeo D em relação ao (2R,3R)-pterosídeo C. As interações de acoplamento da (2R,3R)-pterosina C apresentadas na Fig. 4d mostram o grupo 3-OH do anel indanone ligado a Ser36 e Asn37 através de duas ligações de hidrogênio a distâncias de 1,83 e 2,07 Å, respectivamente. Ala39, Val69, Trp76, Ile118 e Arg128 estiveram envolvidos em interações hidrofóbicas, enquanto Val69 exibiu uma interação π-sigma.
As poses acopladas de (2R)-pterosídeo B, (2S,3R)-pterosina C, e (2R)-pterosina B (inibidores não competitivos do BACE1) são mostradas na Fig. 4e-g, respectivamente. Eles foram acoplados na cavidade fechada por Asn37, Val69, Tyr71, Trp76, Lys107, Phe108, e Ile126. De acordo com seus valores de atividade, (2R)-pterosídeo B (IC50 = 18,0 µM), (2S,3R)-pterosina C (IC50 = 23,1 µM), e (2R)-pterosina B (IC50 = 29,6 µM) exibiram um B.E. de -6,16, -5,07, e -4,64 kcal/mol, respectivamente. Como mostrado na Fig. 4e, (2R)-pterosídeo B demonstrou maior atividade do que (2S,3R)-pterosina C e (2R)-pterosina B devido à presença de um grupo adicional de 2-hidroximetil-tetra-hidro-pirano-3,4,5-triol, que mostrou quatro interações de ligação de hidrogênio. Foram observadas duas ligações de hidrogênio entre o grupo 4-OH do anel tetrahidro-pirano-triol e os grupos NH e CO de Asn37, a distâncias de 2,48 e 2,19 Å, respectivamente. Além disso, os grupos 3 e 5-OH apresentaram duas ligações adicionais de hidrogênio com Ile126 e Trp76 a distâncias de 2,19 e 2,46 Å, respectivamente. (2S,3R)-Pterosina C mostrou uma atividade ligeiramente melhor que (2R)-pterosina B devido à presença de um grupo OH adicional na posição 3 do anel indanônico, que formou uma ligação de hidrogênio com Lys107 a uma distância de 2,12 Å (Fig. 4f). As outras interações foram similares às da (2R)-pterosina B. Como mostrado na Fig. 4g, (2R)-pterosina B mostrou interações hidrofóbicas com Val69, Tyr76, e Phe108.
ACE docking
(2R)-Pteróside B e (2R,3R)-pterosina C foram selecionados como representantes para demonstrar os modos acoplados de inibidores de AChE mistos e não competitivos, respectivamente, devido às suas atividades e tipo de inibição de AChE. A Figura 5a, b ilustra os modelos acoplados de (2R)-pterosídeo B e (2R,3R)-pterosina C, respectivamente. As interações dos compostos acoplados dentro do sítio ativo do AChE são mostradas na Fig. 6.

Modelos de acoplamento molecular para o inibidor misto de AChE (2R)-pterosídeo B (cor azul) e b o inibidor não competitivo de AChE (2R,3R)-pterosina C (cor marrom). Posições bloqueadas são sobrepostas à estrutura de cristal de raios X do E2020 (cor amarela) (código PDB: 4EY7). AChE, resíduos ativos do local e compostos são mostrados por modelos de fita, linha e bastão, respectivamente. As cores das linhas pontilhadas explicam os tipos de várias interacções: interacções de ligação de hidrogénio (verde) e interacções hidrofóbicas (cor-de-rosa). Acetilcolinesterase AChE

Diagrama de interação ligand de um E2020, b (2R)-pterosídeo B, c (2R,3R)-pterosídeo C, d (2S,3R)-pterosina C, e (2R)-pterosina B, f (3S)-pterosídeo D e g (2R,3R)-pterosina C no sítio ativo do AChE. As cores das linhas pontilhadas explicam os tipos de várias interacções: interacções de ligação ao hidrogénio (verde), interacções hidrofóbicas (rosa) e interacções π-sigma (púrpura). AChE acetilcolinesterase
A posição acoplada do E2020 demonstrou um B.E. de -10,28 kcal/mol. Como ilustrado na Fig. 6a, o grupo CO do anel indanônico formou uma ligação de hidrogênio com o grupo NH de Phe295 a uma distância de 1,70 Å. Trp286 e Tyr341 estiveram envolvidos em interações π-sigma, enquanto que Trp86, Trp286, Tyr337, Phe338 e Tyr341 mediaram interações hidrofóbicas. A Figura 6b-f demonstra as poses acopladas de (2R)-pterosídeo B, (2R,3R)-pterosídeo C, (2S,3R)-pterosídeo C, (2R)-pterosídeo B e (3S)-pterosídeo D (inibidores de AChE de tipo misto), respectivamente. Eles foram acomodados no local ativo cercados por Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338, e Tyr341. Consistente com seus valores de atividade, (2R)-pterosídeo B (IC50 = 2,55 µM), (2R,3R)-pterosídeo C (IC50 = 3,77 µM), (2S,3R)-pterosídeo C (IC50 = 12.8 µM), (2R)-pterosina B (IC50 = 16,2 µM) e (3S)-pterosídeo D (IC50 = 27,4 µM) exibiram um B.E. de -7,90, -7,49, -6,03, -5,76, e -4,91 kcal/mol, respectivamente. (2R)-Pterósido B demonstrou uma potência maior que (2S,3R)-pterosina C e (2R)-pterosina B devido à presença de um grupo adicional de 2-hidroximetil-tetra-hidro-pirano-3,4,5-triol, que estabeleceu três interações de ligação de hidrogênio (Fig. 6b). O grupo OH de hidroximetil e o grupo 3-OH do anel tetrahidro-piranotriol apresentaram ligações de hidrogênio com o grupo CO de Asn87 a distâncias de 2,23 e 2,28 Å, respectivamente. Além disso, o grupo 4-OH formou uma ligação de hidrogênio com o grupo CO do Trp86 a uma distância de 2,13 Å. No caso do (2R,3R)-pterosídeo C (Fig. 6c), o grupo metilo na posição 2 do anel de indanona não apresentou interação hidrofóbica com o Tyr341 e, portanto, exibiu atividade comparativamente menor que (2R)-pterosídeo B. Entretanto, Tyr341 manteve a interação hidrofóbica com a outra parte do anel de indanona como mostrado na do (2R)-pterosídeo B. A maior atividade de (2S,3R)-pterosina C que (2R)-pterosina B foi atribuída à existência de um grupo OH adicional na posição-3 do anel de indanona, que formou uma ligação de hidrogênio com Phe295 a uma distância de 1,82 Å (Fig. 6d). As demais interações foram comparáveis às interações (2R)-pterosina B. Como mostrado na Fig. 6e, o grupo OH do grupo hidroxietil na posição-6 do anel de indanona formou uma ligação de hidrogênio com Tyr124 a uma distância de 2,48 Å. Tyr124, Trp286, Tyr337, Phe338, e Tyr341 contribuíram para as interações hidrofóbicas. Em comparação com (2R)-pterosídeo B e (2R,3R)-pterosídeo C, (3S)-pterosídeo D apresentaram interações de ligação dissimilares devido à presença do grupo 2,2-dimetil no anel de indanona (Fig. 6f). O grupo 2,2-dimetil contribuiu significativamente para a distinta posição acoplada do (3S)-pterosídeo D. O grupo 5-OH do anel tetrahidro-pirano-triol formou uma ligação de hidrogênio com Asp74 a uma distância de 1,96 Å. O grupo CO do anel de indanona mostrou uma ligação de hidrogênio com Tyr72 a uma distância de 2,97 Å. Essas interações foram responsáveis pela baixa atividade do (3S)-pterosídeo D.
A pose acoplada de (2R,3R)-pterosina C (inibidor AChE não competitivo) é exibida na Fig. 6g. (2R,3R)-Pterosina C (IC50 = 23,2 µM) demonstrou um B.E. de -5,01 kcal/mol. A bolsa de ligação de (2R,3R)-pterosina C compreendeu Tyr72, Trp286, Ser293, Phe297, e Tyr341, com duas interações de ligação de hidrogênio. Uma das ligações de hidrogênio foi formada entre o grupo 3-OH do anel de indanona e o grupo CO do Ser293 a uma distância de 2,03 Å. A segunda ligação de hidrogênio foi observada entre o grupo OH do grupo hidroxietil presente na posição-6 do anel de indanona e Tyr72 a uma distância de 1,94 Å. Resíduos como Tyr72, Trp286, Phe297 e Tyr341 participaram de interações hidrofóbicas.
Acoplamento de BChE
Considerando os níveis de atividade e o tipo de inibição de BChE, (2R,3R)-pterosídeo C e (2S,3R)-pterosina C foram selecionados para demonstrar os modos acoplados de inibidores de BChE do tipo misto e não-competitivo, respectivamente. A Figura 7a, b ilustra os modelos acoplados de (2R,3R)-pterosídeo C e (2S,3R)-pterosina C, respectivamente. As interações dos compostos acoplados dentro do sítio ativo do BChE são apresentadas na Fig. 8.
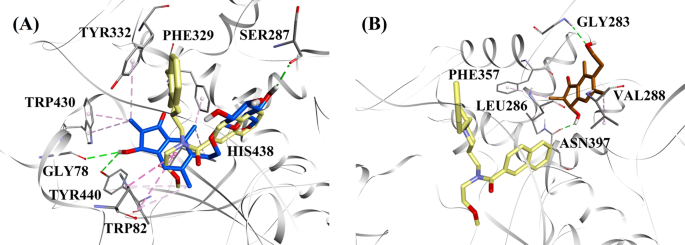
Modelos de acoplamento molecular para o inibidor misto BChE (2R,3R)-pterosídeo C (cor azul) e b o inibidor não competitivo BChE (2S,3R)-pterosina C (cor marrom). Posições bloqueadas são sobrepostas na estrutura de cristal de raio X de 3F9 (cor amarela) (código PDB: 4TPK). BChE, resíduos e compostos ativos do local são mostrados por modelos de fita, linha e bastão, respectivamente. As cores das linhas pontilhadas explicam os tipos de várias interações: interações de ligação de hidrogênio (verde) e interações hidrofóbicas (rosa). Butilcolinesterase BChE
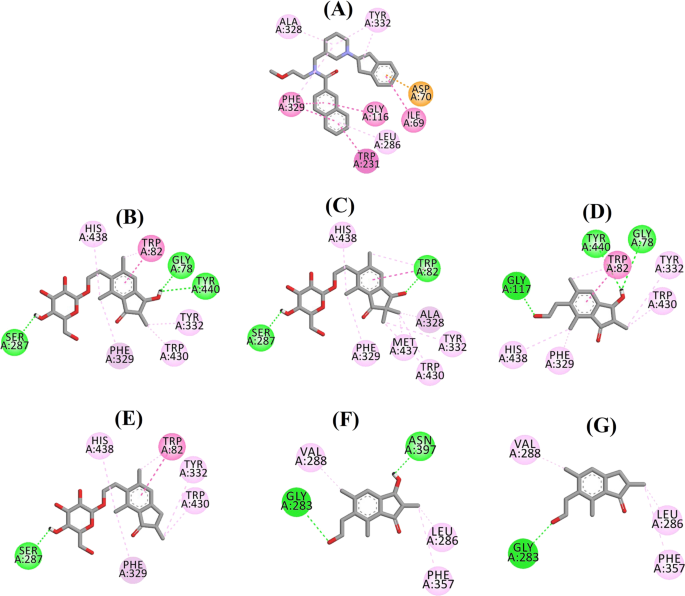
Diagrama de interação ligand de um 3F9, b (2R,3R)-pterosídeo C, c (3S)-pterosídeo D, d (2R,3R)-pterosídeo C, e (2R)-pterosídeo B, f (2S,3R)-pterosídeo C e g (2R)-pterosídeo B no site ativo do BChE. As cores das linhas pontilhadas explicam os tipos de várias interações: interações de ligação de hidrogênio (verde), interações hidrófobas (rosa) e interações de aniões π (dourado). BChE butyrylcholinesterase
A posição acoplada de 3F9 mostrou um B.E. de -8,49 kcal/mol. Como mostrado na Fig. 8a, as interações hidrofóbicas foram as principais responsáveis pela ligação ligand. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329, e Tyr332 foram responsáveis pelas interações hidrofóbicas, enquanto Asp70 demonstrou uma interação π-anion. A Figura 8b-e ilustra as poses acopladas de (2R,3R)-pterosídeo C, (3S)-pterosídeo D, (2R,3R)-pterosídeo C e (2R)-pterosídeo B (inibidores BChE de tipo misto), respectivamente. Sua bolsa de ligação foi composta de Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438, e Tyr440. De acordo com seus níveis de atividade, (2R,3R)-pterosídeo C (IC50 = 5,29 µM), (3S)-pterosídeo D (IC50 = 19,3 µM), (2R,3R)-pterosídeo C (IC50 = 20,3 µM) e (2R)-pterosídeo B (IC50 = 62,0 µM) demonstraram um B.E. de -7,23, -6,59, -6,52, e -4,38 kcal/mol, respectivamente. Como mostrado na Fig. 8b, o grupo 3-OH do anel indanone de (2R,3R)-pterosídeo C mostrou duas ligações de hidrogênio com Gly78 e Tyr440 em distâncias de 2,87 e 2,89 Å, respectivamente. No caso do grupo (3S)-pterosídeo D, o grupo 2,2-dimetil no anel de indanona afetou as interações de ligação do grupo 3-OH (Fig. 8c). O grupo 3-OH formou apenas uma ligação de hidrogênio com Trp82 a uma distância de 2,94 Å, o que resultou em uma atividade comparativamente menor de (3S)-pterosídeo D do que (2R,3R)-pterosídeo C. Como mostrado na Fig. 8d, a (2R,3R)-pterosina C falhou em produzir uma ligação de hidrogênio com Ser287 devido à ausência de um grupo 2-hidroximetil-tetra-hidro-pirano-3,4,5-triol. Consequentemente, produziu uma atividade menor que (2R,3R)-pterosídeo C. A falta do grupo OH na posição-3 do anel indanone foi considerada responsável pela atividade muito baixa do (2R)-pterosídeo B (Fig. 8e), que falhou em formar ligações de hidrogênio com Gly78 e Tyr440 semelhantes a (2R,3R)-pterosídeo C.
As poses acopladas de (2S,3R)-pterosina C e (2R)-pterosina B (inibidores não competitivos de BChE) são mostradas na Fig. 8f, g, respectivamente. Essas poses acopladas foram contidas na cavidade fechada por Gly283, Leu286, Val288, Phe357, e Asn397. Conforme seus níveis de atividade, (2S,3R)-pterosina C (IC50 = 44,3 µM) e (2R)-pterosina B (IC50 = 48,1 µM) exibiram um B.E. de -5,40 e -5,06 kcal/mol, respectivamente. (2S,3 R)-Pterosina C foi mais potente que (2R)-pterosina B devido à presença de um grupo OH adicional na posição-3 do anel indanônico, que formou uma ligação de hidrogênio com o grupo CO de Asn397 a uma distância de 2,05 Å (Fig. 8f). Outras interações foram semelhantes à (2R)-pterosina B. Como mostrado na Fig. 8g, o grupo OH no grupo hidroxietil na posição-6 do anel de indanona formou uma ligação de hidrogênio com Gly283 a uma distância de 2,40 Å. Os resíduos Leu286, Val288 e Phe357 participaram de interações hidrofóbicas.
Inibidores do tipo mistura ligam-se tanto à enzima livre quanto ao complexo enzimático-substrato, o que indica que esses compostos podem se ligar ao sítio catalítico de cada enzima correspondente. Os inibidores não-competitivos ligam-se ao sítio alostárico da enzima livre ou do complexo enzimático-substrato. Um estudo recente sugeriu que inibidores competitivos, de tipo misto e não competitivos ocupam locais diferentes nas bolsas de ligação de BACE1, AChE, e BChE38. Durante o docking para avaliação do mecanismo inibitório dos derivados de pterosina, os sítios de ligação dos compostos foram definidos de acordo com o seu tipo de inibição. Os resultados do docking indicaram que os sítios de ligação dos inibidores mistos e não competitivos para BACE1, AChE e BChE se sobrepõem parcialmente em cada sítio ativo correspondente e foram consistentes com um relatório anterior38,
Permeabilidade do BBC
PAMPA-BBBB, um ensaio in vitro de permeabilidade de membrana artificial para o BBB, é uma das ferramentas de triagem físico-química mais confiáveis no estágio inicial da descoberta de drogas direcionadas ao SNC40. O sistema PAMPA-BBB modela a difusão passiva transcelular de produtos químicos através do BBB e mede estritamente mecanismos de transporte passivos através de uma membrana lipídica artificial sobre a permeabilidade efetiva (Pe, cm/s). Com base no padrão estabelecido para a previsão da permeabilidade BBB, os compostos foram classificados em (i) “CNS+” (alta permeação BBB prevista); Pe (10-6 cm/s) > 4,00, (ii) “CNS-” (baixa permeação BBB prevista); Pe (10-6 cm/s) < 2,00, e (iii) “CNS+/-” (permeação BBB incerta); Pe (10-6 cm/s) das 4,00 às 2,00. Assim, (2R)-pterosina B, (2S)-pterosina P, e (2S)-pterosina A exibiram alta permeação BBB com valores Pe de 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s, e 6,26 × 10-6 cm/s, respectivamente (Tabela 4, Informações Suplementares 4). O valor de Pe da (2R)-pterosina B foi 1,7 vezes maior que o do verapamil da droga CNS (Pe = 34,6 × 10-6 cm/s), que foi usado para o controle positivo. (2S,3R)-Pterosina C e (2R,3R)-pterosina C apresentaram permeação incerta do BBB com valores de Pe de 2,34 e 1,98, respectivamente. (2R,3R)-Pterosida C, (3S)-pterosida D e (2R)-pterosida B, que mostraram as mais potentes atividades inibitórias de BACE1 e colinesterase entre os derivados de pterosina testados, exibiram uma permeabilidade muito baixa do BBB. A existência do grupo 2-hidroximetil-tetra-hidro-pirano-3,4,5-triol como nos derivados pterósidos resultou em uma notável diminuição da permeabilidade ao BBB. Em comparação com a (2R)-pterosina B, a presença adicional do grupo OH na posição-3 do anel de indanona como na pterosina C, o grupo hidroximetil na posição-2 do anel de indanona como na (2S)-pterosina A ou o grupo hidroximetil na posição-5 do anel de indanona como na (2S)-pterosina P também reduziu significativamente a permeabilidade do BBB. Considerando uma permeabilidade excepcionalmente alta do BBB e a significativa inibição do BACE1, AChE e BChE, (2R)-pterosina B pode ter o potencial de exibir uma forte atividade anti-AD.
Efeitos da (2R)-pterosina B e (2R,3R)-pterosídeo C na secreção de Aβ peptídeos por células neuronais
Investigar a função da (2R)-pterosina B e (2R)-pterosina C,3R)-pterosídeo C ao diminuir a excreção de Aβ das células neuronais, utilizamos uma linha de células do neuroblastoma murino que, de forma estável, expressa excessivamente o APPswe humano. A linha de células é um modelo celular de AD caracterizado pela secreção excessiva de Aβ40 e Aβ42. Os oligômeros amilóides tóxicos são formados a partir das duas isoformas do peptídeo Aβ com diferentes comprimentos. Aβ40 é a isoforma Aβ mais abundante no cérebro41, enquanto Aβ42 aumenta significativamente com certas formas de AD42. O Sandwich ELISA de Aβ40 mostrou que a (2R)-pterosina B reduziu significativamente a quantidade de peptídeo Aβ40 secretado das células do neuroblastoma em meio até 50% a 500 μM (P < 0,01) (Fig. 9a). Da mesma forma, a secreção de Aβ42 peptídeo pelas células do neuroblastoma diminuiu significativamente na presença de 500 µM de (2R,3R)-pterosídeo C (P < 0,05) (Fig. 9b). Em conclusão, (2R)-pterosina B e (2R,3R)-pterosídeo C diminuíram significativamente a secreção de peptídeos de células do neuroblastoma Aβ a uma concentração de 500 μM.

a Efeito da (2R)-pterosina B na secreção dos peptídeos do Aβ. As células do neuroblastoma secretor de APPswe foram tratadas com 12, 60, 250 ou 500 µM de (2R)-pterosina B durante 24 h, e meios condicionados foram coletados na presença do inibidor de protease. β-SI (10 µM) foi usado como controle positivo. O controle negativo incluiu células cultivadas na ausência de compostos de teste. A análise quantitativa da secreção de Aβ40 e Aβ42 na mídia condicionada foi realizada utilizando o ELISA em sanduíche. As médias ± SD de três experimentos independentes são mostradas. Os peptídeos secretados Aβ40 diminuíram significativamente na presença de 500 µM (2R)-pterosina B. CON controle negativo, β-SI β- inibidor de secreção III, Aβ β-amilóide. *P < 0,05, **P < 0,01. b Efeito do (2R,3R)-pterosídeo C sobre a secreção de Aβ peptídeos. As células do neuroblastoma secretor de (2R,3R)-pterosídeo C foram tratadas com 12, 60, 125, 250, ou 500 µM de (2R,3R)-pterosídeo C durante 24 h, e meios condicionados foram coletados na presença de inibidor de protease. β-SI (10 µM) foi usado como controle positivo. O controle negativo incluiu células cultivadas na ausência de compostos de teste. A análise quantitativa do secreto Aβ40 e Aβ42 no meio condicionado foi realizada utilizando o sanduíche ELISA. São mostradas as médias ± SD de três experimentos independentes. O peptídeo secretado Aβ42 diminuiu significativamente na presença de 500 µM (2R,3R)-pterosídeo C. CON controle negativo, β-SI β- inibidor de secreção III, Aβ β-amilóide. *P < 0.05
Citotoxicidade dos derivados de pterosina com base no ensaio MTT43
Os derivados de pterosina, em geral, apresentaram citotoxicidade desprezível contra várias linhas celulares normais e cancerígenas, como o neuroblastoma humano SH-SY5Y, células gliais de rato C6, fibroblastos embrionários de rato NIH3T3 e melanoma de rato B16F10 com valores de LD50 acima de 0.5 mM (Informação suplementar 5). Em particular, a (2R)-pterosina B, (3R)-pterosina D, (2S)-pterosina P, (2S)-pterosídeo A, (2R)-pterosídeo B, e (2R,3R)-pterosídeo C não apresentaram citotoxicidade contra as linhas celulares testadas com valores de LD50 acima de 5 mM. Intrigantemente, várias pterosinas mostraram efeitos antiproliferativos relativos contra as células neuronais SH-SY5Y em comparação com as células gliais C6 de origem mesenquimatosa. Os resultados presentes foram consistentes com um relatório anterior que sugeria que os derivados de pterosina são atóxicos para humanos26,