

>
>
A contribuição de uma ponte salina para a estabilidade geral ao estado dobrado de uma proteína pode ser avaliada através de dados termodinâmicos coletados de estudos de mutagênese e técnicas de ressonância magnética nuclear. Usando uma proteína do tipo pseudo-mutativa, especificamente mutada para prevenir a precipitação a pH elevado, a contribuição da ponte salina para a energia livre global do estado de proteína dobrada pode ser determinada pela realização de uma mutação pontual, alterando e, consequentemente, quebrando a ponte salina. Por exemplo, foi identificada a existência de uma ponte salina na lisozima T4 entre o ácido aspártico (Asp) no resíduo 70 e uma histidina (His) no resíduo 31 (Figura 3). A mutagénese local com asparagina (Asn) (Figura 4) foi feita obtendo três novos mutantes: Asp70Asn His31 (Mutante 1), Asp70 His31Asn (Mutante 2) e Asp70Asn His31Asn (Mutante Duplo).

>
>
Uma vez estabelecidos os mutantes, dois métodos podem ser empregados para calcular a energia livre associada a uma ponte de sal. Um método envolve a observação da temperatura de fusão da proteína do tipo selvagem versus a dos três mutantes. A desnaturação pode ser monitorada através de uma mudança no dicroísmo circular. Uma redução na temperatura de fusão indica uma redução na estabilidade. Isto é quantificado através de um método descrito por Becktel e Schellman onde a diferença de energia livre entre os dois é calculada através de ΔTΔS. Há alguns problemas com este cálculo e só pode ser usado com dados muito precisos. No exemplo da lisozima T4, ΔS do tipo pseudo-molha tinha sido previamente reportado a pH 5,5, portanto a diferença de temperatura média de 11 °C a este pH multiplicada pelo reportado ΔS de 360 cal/(mol-K) (1,5 kJ/(mol-K)) produz uma mudança de energia livre de cerca de -4 kcal/mol (-17 kJ/mol). Este valor corresponde à quantidade de energia livre que contribui para a estabilidade da proteína pela ponte salina.
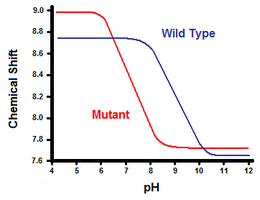
O segundo método utiliza espectroscopia de ressonância magnética nuclear para calcular a energia livre da ponte de sal. Uma titulação é realizada, enquanto se registra o deslocamento químico correspondente aos prótons do carbono adjacente ao grupo do carboxilato ou amônio. O ponto médio da curva de titulação corresponde ao pKa, ou o pH onde a proporção de moléculas protonadas:desprotonadas é de 1:1. Continuando com o exemplo da lisozima T4, uma curva de titulação é obtida através da observação de uma mudança no próton C2 de histidina 31 (Figura 5). A Figura 5 mostra o deslocamento da curva de titulação entre o tipo selvagem e o mutante no qual Asp70 é Asn. A ponte de sal formada é entre a Asp70 desprotonada e a His31 protonada. Esta interação causa o deslocamento visto na pKa da His31. Na proteína do tipo selvagem desdobrada, onde a ponte de sal está ausente, a His31 é relatada como tendo uma pKa de 6,8 em tampões H20 de força iônica moderada. A Figura 5 mostra uma pKa do tipo selvagem de 9,05. Esta diferença em pKa é suportada pela interação do His31 com o Asp70. Para manter a ponte de sal, o His31 tentará manter o protão o máximo de tempo possível. Quando a ponte de sal é interrompida, como na D70N mutante, o pKa volta a um valor de 6,9, muito mais próximo ao de His31 no estado desdobrado.
A diferença em pKa pode ser quantificada para refletir a contribuição da ponte de sal para a energia livre. Usando a energia livre de Gibbs:ΔG = -RT ln(Keq), onde R é a constante universal do gás, T é a temperatura em kelvins, e Keq é a constante de equilíbrio de uma reação em equilíbrio. A desprotonação de His31 é uma reação de equilíbrio ácido com uma Keq especial conhecida como a constante de dissociação ácida, Ka: His31-H+ ⇌ His31 + H+. A pKa é então relacionada a Ka pelo seguinte: pKa = -log(Ka). O cálculo da diferença de energia livre do tipo mutante e selvagem pode agora ser feito usando a equação de energia livre, a definição de pKa, os valores de pKa observados e a relação entre logaritmos naturais e logaritmos. No exemplo da lisozima T4, esta abordagem produziu uma contribuição calculada de cerca de 3 kcal/mol para a energia livre global. Uma abordagem semelhante pode ser feita com o outro participante na ponte de sal, como Asp70 no exemplo da lisozima T4, monitorando seu deslocamento em pKa após a mutação de His31.
Uma palavra de precaução ao escolher a experiência apropriada envolve a localização da ponte de sal dentro da proteína. O ambiente desempenha um grande papel na interação. Em altas forças iônicas, a ponte de sal pode ser completamente mascarada, uma vez que uma interação eletrostática está envolvida. A ponte de sal His31-Asp70 em lisozima T4 foi enterrada dentro da proteína. A entropia desempenha um papel maior nas pontes salinas de superfície onde os resíduos que normalmente têm a capacidade de se moverem são apertados pela sua interacção electrostática e ligação de hidrogénio. Isto mostrou diminuir a entropia o suficiente para quase apagar a contribuição da interação. As pontes salinas superficiais podem ser estudadas de forma semelhante à das pontes salinas enterradas, empregando ciclos duplos mutantes e titulações NMR. Embora existam casos em que pontes salinas enterradas contribuem para a estabilidade, como qualquer outra coisa, existem excepções e as pontes salinas enterradas podem mostrar um efeito desestabilizador. Além disso, as pontes salinas superficiais, sob certas condições, podem exibir um efeito estabilizador. O efeito estabilizador ou desestabilizante deve ser avaliado caso a caso e poucas afirmações sobre a camada podem ser feitas.