

Bidraget från en saltbrygga till den totala stabiliteten till ett proteins veckade tillstånd kan bedömas med hjälp av termodynamiska data som samlats in från mutagenesestudier och kärnmagnetresonanstekniker. Med hjälp av ett muterat protein av pseudovildtyp som är specifikt muterat för att förhindra utfällning vid högt pH kan saltbryggans bidrag till den totala fria energin i det veckade proteintillståndet bestämmas genom att utföra en punktmutation som ändrar och följaktligen bryter saltbryggan. Till exempel identifierades en saltbrygga i T4-lysozym mellan asparaginsyra (Asp) vid rest 70 och ett histidin (His) vid rest 31 (figur 3). Genom platsriktad mutagenes med asparagin (Asn) (figur 4) erhölls tre nya mutanter: Asp70Asn His31 (mutant 1), Asp70 His31Asn (mutant 2) och Asp70Asn His31Asn (dubbelmutant).

När mutanterna har fastställts kan två metoder användas för att beräkna den fria energin i samband med en saltbrygga. Den ena metoden innebär att man observerar smälttemperaturen för vildtypsproteinet jämfört med smälttemperaturen för de tre mutanterna. Denatureringen kan övervakas genom en förändring i cirkulär dikroism. En sänkning av smältningstemperaturen tyder på minskad stabilitet. Detta kvantifieras genom en metod som beskrivs av Becktel och Schellman där den fria energidifferensen mellan de två beräknas genom ΔTΔS. Det finns vissa problem med denna beräkning och den kan endast användas med mycket exakta data. I exemplet med T4-lysozym hade ΔS av pseudovildtyp tidigare rapporterats vid pH 5,5, så temperaturskillnaden vid mittpunkten på 11 °C vid detta pH multiplicerat med den rapporterade ΔS på 360 cal/(mol-K) (1,5 kJ/(mol-K)) ger en förändring av den fria energin på cirka -4 kcal/mol (-17 kJ/mol). Detta värde motsvarar den mängd fri energi som saltbryggan bidrar till proteinets stabilitet.
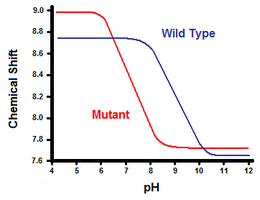
Den andra metoden använder kärnmagnetisk resonansspektroskopi för att beräkna saltbryggans fria energi. En titrering utförs samtidigt som man registrerar det kemiska skiftet som motsvarar protonerna hos kolet intill karboxylat- eller ammoniumgruppen. Mittpunkten på titreringskurvan motsvarar pKa, eller det pH-värde där förhållandet mellan protonerade och deprotonerade molekyler är 1:1. Om vi fortsätter med exemplet med T4-lysozym erhålls en titreringskurva genom observation av en förskjutning av C2-protonen i histidin 31 (figur 5). Figur 5 visar förskjutningen i titreringskurvan mellan vildtypen och mutanten där Asp70 är Asn. Den saltbrygga som bildas är mellan den deprotonerade Asp70 och den protonerade His31. Denna interaktion orsakar den förskjutning som ses i His31:s pKa. I det oveckade vildtypsproteinet, där saltbryggan saknas, rapporteras His31 ha en pKa på 6,8 i H20-buffertar med måttlig jonstyrka. Figur 5 visar en pKa för vildtypen på 9,05. Denna skillnad i pKa stöds av His31:s interaktion med Asp70. För att upprätthålla saltbryggan kommer His31 att försöka behålla sin proton så länge som möjligt. När saltbryggan bryts, som i mutanten D70N, förskjuts pKa tillbaka till ett värde på 6,9, mycket närmare His31:s värde i det utvikta tillståndet.
Den skillnaden i pKa kan kvantifieras för att återspegla saltbryggans bidrag till den fria energin. Med hjälp av Gibbs fria energi: ΔG = -RT ln(Keq), där R är den universella gaskonstanten, T är temperaturen i kelvin och Keq är jämviktskonstanten för en reaktion i jämvikt. Deprotoneringen av His31 är en sur jämviktsreaktion med en speciell Keq som kallas syredissociationskonstanten Ka: His31-H+ ⇌ His31 + H+. pKa relateras sedan till Ka enligt följande: pKa = -log(Ka). Beräkningen av skillnaden i fri energi mellan mutanten och vildtypen kan nu göras med hjälp av ekvationen för fri energi, definitionen av pKa, de observerade pKa-värdena och förhållandet mellan naturliga logaritmer och logaritmer. I exemplet med T4-lysozym gav detta tillvägagångssätt ett beräknat bidrag på cirka 3 kcal/mol till den totala fria energin. Ett liknande tillvägagångssätt kan användas med den andra deltagaren i saltbryggan, t.ex. Asp70 i T4-lysozym-exemplet, genom att övervaka dess förskjutning i pKa efter mutation av His31.
En försiktighetsåtgärd vid val av lämpligt experiment gäller saltbryggans placering i proteinet. Miljön spelar en stor roll för interaktionen. Vid höga jonstyrkor kan saltbryggan vara helt maskerad eftersom en elektrostatisk interaktion är inblandad. Saltbryggan His31-Asp70 i T4 lysozym var begravd i proteinet. Entropi spelar en större roll i ytliga saltbryggor där rester som normalt har möjlighet att röra sig begränsas av sin elektrostatiska interaktion och vätebindning. Detta har visat sig minska entropin tillräckligt mycket för att nästan utplåna interaktionens bidrag. Ytliga saltbryggor kan studeras på samma sätt som begravda saltbryggor, med hjälp av dubbla mutantcykler och NMR-titreringar. Även om det finns fall där begravda saltbryggor bidrar till stabiliteten, finns det precis som med allt annat undantag och begravda saltbryggor kan ha en destabiliserande effekt. Dessutom kan ytliga saltbryggor under vissa förhållanden ha en stabiliserande effekt. Den stabiliserande eller destabiliserande effekten måste bedömas från fall till fall och få generella uttalanden kan göras.