
Isolamento e caratterizzazione dei derivati della pterosina da P. aquilinum
Derivati della pterosina sono stati isolati dalle piante intere di P. aquilinum. Per lo studio dei costituenti fitochimici della felce di salice, un estratto d’acqua è stato successivamente partizionato con acetato di etile (EtOAc) e n-butanolo (n-BuOH) (Fig. 1). La cromatografia su colonna ripetuta della frazione solubile in EtOAc ha portato all’isolamento di nove derivati, tra cui (2R)-pterosina B (1), pterosina Z (2), (2S)-pterosina P (3), (3R)-pterosina D (4), (2S)-pterosina A (5), (2S,3R)-pterosina C (6), (2R,3R)-pterosina C (7), (2R)-pteroside B (8), e pteroside Z (9), con purezza superiore al 97% (Fig. 2). La ripetuta separazione cromatografica su colonna della frazione solubile in n-BuOH ha portato all’isolamento di sei derivati, tra cui (2S)-pteroside A (10), (2S)-pteroside A2 (11), (2S,3R)-pteroside C (12), (2R,3R)-pteroside C (13), (3S)-pteroside D (14), e (2S)-pteroside P (15), con purezze superiori al 97%. Le strutture dei composti sono state identificate dall’analisi dei dati spettrali, tra cui MS, 1D- e 2D-NMR (Informazioni supplementari 1).

Strutture dei composti di pterosina 1-15xxx
Attività inibitoria dei derivati di pterosina contro BACE1, AChE, e BChE
Per valutare il potenziale anti-AD, l’attività inibitoria di ogni composto della pterosina contro BACE1 e colinesterasi è stata valutata mediante i rispettivi saggi di inibizione in vitro (Tabella 1). Tutti i derivati della pterosina testati hanno mostrato attività inibitoria concentrazione-dipendente contro BACE1 con un range di valori IC50 (concentrazione inibitoria semimassima) di 9,74-94,4 μM, con l’eccezione di (2S)-pterosina A e (2S)-pteroside P che erano inattivi alle concentrazioni testate, rispetto all’IC50 della quercetina usata come controllo positivo, che era 18,8 μM. La potenza inibitoria degli inibitori più forti era nell’ordine di (2R,3R)-pteroside C, (3S)-pteroside D, (2R)-pteroside B, (2S,3R)-pterosina C, (2R,3R)-pterosina C, (2S,3R)-pteroside C e (2R)-pterosina B con valori IC50 di 9.74, 10.7, 18.0, 23.1, 26.2, 28.9, e 29.6 μM, rispettivamente. Abbiamo successivamente testato i potenziali inibitori dei derivati della pterosina contro AChE. Tutti i composti testati hanno mostrato significative attività AChE-inibitorie, con valori IC50 nell’intervallo di 2,55-110 μM, rispetto all’IC50 contro AChE della berberina usata come controllo positivo, che era 0,39 μM. I composti di pterosina che hanno mostrato la più forte attività inibitoria contro AChE erano (2R)-pteroside B, (2R,3R)-pteroside C, (2S,3R)-pteroside C, (2S,3R)-pterosina C, e (2R)-pterosina B con valori IC50 di 2,55, 3,77, 9,17, 12,8, e 16,2 μM, rispettivamente. Infine, abbiamo testato le capacità inibitorie dei derivati della pterosina contro BChE. Tutti i composti di pterosina testati hanno mostrato attività inibitoria contro BChE, con valori IC50 che variavano da 5,29 a 119 μM, con l’eccezione di (3R)-pterosina D che era inattiva alle concentrazioni testate, rispetto all’IC50 della berberina contro BChE, che era 3,32 μM. I composti di pterosina che hanno mostrato la più forte attività inibitoria contro BChE sono stati (2R,3R)-pteroside C e pteroside Z con valori IC50 di 5,29 e 5,31 μM, rispettivamente.
Collettivamente, la maggior parte dei derivati della pterosina testati ha mostrato attività inibitorie significative contro BACE1, AChE e BChE contemporaneamente. La presenza del gruppo aggiuntivo 2-idrossimetil-tetraidro-pirano-3,4,5-triolo come nei derivati della pteroside ha aumentato significativamente le attività inibitorie contro gli enzimi. Inoltre, la presenza del gruppo idrossimetilico aggiuntivo in posizione-2 dell’anello indanonico della (2R)-pterosina B come nella (2S)-pterosina A o il gruppo metilico come nella (3R)-pterosina D e nella pterosina Z ha diminuito le attività inibitorie contro gli enzimi. In particolare, la presenza del gruppo idrossimetilico in posizione 5 dell’anello indanonico come nella (2S)-pterosina P diminuiva l’attività inibitoria contro BACE1.
Parametri cinetici dell’inibizione enzimatica da parte dei derivati della pterosina
Nel tentativo di spiegare la modalità di inibizione enzimatica dei derivati della pterosina, abbiamo eseguito un’analisi cinetica di BACE1 e colinesterasi per gli inibitori rappresentativi (Tabella 2, Informazioni supplementari 2). Un basso Ki (costante di inibizione) indica un legame enzimatico più stretto e un inibitore più efficace. Nel complesso, i valori Ki dei composti erano correlati ai rispettivi valori IC50. L’inibizione di BACE1 da parte dei composti (2R,3R)-pteroside C, (3S)-pteroside D, e (2R,3R)-pterosina C era di tipo misto con valori Ki di 12,6, 16,5, e 27.6 µM, rispettivamente, mentre l’inibizione da (2R)-pteroside B, (2S,3R)-pteroside C, e (2R)-pterosina B era non competitiva con valori Ki di 23,1, 33,8, e 38,3 µM, rispettivamente. L’inibizione di AChE da parte di (2R)-pteroside B, (2R,3R)-pteroside C, (2R)-pterosina B, (2S,3R)-pterosina C, e (3S)-pteroside D era di tipo misto con valori Ki di 4.89, 8,13, 12,1, 16,3 e 23,1 µM, rispettivamente, mentre la (2R,3R)-pterosina C era un inibitore di tipo non competitivo con un valore Ki di 29,6 µM. L’inibizione di BChE da parte di (2R,3R)-pterosina C, (2R,3R)-pteroside C, (3S)-pteroside D, e (2R)-pteroside B era di tipo misto con valori Ki di 4,77, 9,62, 19.7 e 22,6 µM, rispettivamente, mentre (2S,3R)-pteroside C e (2R)-pteroside B erano inibitori non competitivi con valori Ki di 29,9 e 53,5 µM, rispettivamente. Quindi, questi risultati hanno suggerito che i derivati specifici della pterosina potrebbero essere efficaci inibitori di BACE1, AChE e BChE.
Simulazioni di docking molecolare per BACE1, AChE, e BChE
Sono disponibili diverse strutture cristalline per BACE1 e colinesterasi. Abbiamo selezionato i PDB umani in base alle strutture wild-type, ai ligandi cocristallizzati e alle risoluzioni delle strutture. Le strutture cristalline a raggi X di BACE1 complessato con QUD (codice PDB: 2WJO, risoluzione: 2.5 Å)33, AChE complessato con E2020 (codice PDB: 4EY7, risoluzione: 2.35 Å)34, e BChE complessato con 3F9 (codice PDB: 4TPK, risoluzione: 2.70 Å)35 sono state selezionate per il docking. Inizialmente, QUD, E2020 e 3F9 sono stati estratti dalle strutture cristalline e reinseriti nei siti attivi di BACE1, AChE e BChE, rispettivamente. Successivamente, (2R,3R)-pteroside C, (3S)-pteroside D, (2R,3R)-pteroside C, (2R)-pteroside B, (2S,3R)-pteroside C e (2R)-pteroside B con il noto meccanismo di inibizione contro BACE1, AChE e BChE sono stati inseriti per determinare il loro SAR. I risultati del docking sono riassunti nella tabella 3. Il SAR degli inibitori selezionati di tipo misto e non competitivo di BACE1, AChE e BChE ha permesso di valutare la nuova inibizione diretta al sito di legame degli enzimi da parte delle pterosine.
La nostra modalità di docking di E2020 era coerente con la modalità di legame determinata sperimentalmente e precedentemente riportata con AChE umana ricombinante (rhAChE) (Informazioni supplementari 3)34. La radice media dello scarto quadratico (RMSD) tra il cristallo e le conformazioni agganciate di E2020 era 0,54 Å, che ha suggerito l’affidabilità della nostra configurazione docking nel riprodurre la modalità di legame sperimentale. Inoltre, la modalità docked di E2020 ha portato a un’interazione simile a quella di rhAChE-E2020. Nel nostro studio, le molecole d’acqua sono state rimosse dalla struttura cristallina durante il docking; pertanto, le interazioni mediate dall’acqua non sono state analizzate nel presente studio. Allo stesso modo, le modalità docked di QUD e 3F9 erano coerenti con i dati sperimentali disponibili per BACE1 33 e BChE35, rispettivamente (Informazioni supplementari 3). Le RMSD tra il cristallo e conformazioni docked di QUD e 3F9 erano 0,46 e 0,60 Å, rispettivamente. Inoltre, i siti di legame degli inibitori pterosin erano in accordo con un precedente studio di docking che ha coinvolto BACE1, AChE, e BChE38. Tuttavia, lo studio ha utilizzato Tetronarce californica AChE (codice PDB: 1ACJ), che contiene numeri di residui leggermente diversi da AChE umano a causa di variazioni nelle loro sequenze.
BACE1 docking
In base al tipo di inibizione e attività, (2R,3R)-pteroside C e (2R)-pteroside B sono stati selezionati per dimostrare le modalità docked di tipo misto e non competitivo inibitori BACE1, rispettivamente. Figura 3a, b mostra i modelli di docking di (2R,3R)-pteroside C e (2R)-pteroside B, rispettivamente. Le interazioni dei composti agganciati all’interno del sito attivo di BACE1 sono mostrate in Fig. 4.

Modelli di docking molecolare per a l’inibitore di tipo misto BACE1 (2R,3R)-pteroside C (colore blu) e b l’inibitore non competitivo BACE1 (2R)-pteroside B (colore marrone). Le pose di dock sono sovrapposte alla struttura cristallina a raggi X di QUD (colore giallo) (codice PDB: 2WJO). BACE1, i residui del sito attivo e i composti sono mostrati rispettivamente da modelli a nastro, linea e bastone. I colori delle linee tratteggiate spiegano i tipi di varie interazioni: interazioni di legame idrogeno (verde), interazioni idrofobiche (rosa) e interazioni π-sigma (viola). BACE1 β-sito dell’enzima di scissione della proteina precursore dell’amiloide 1

Diagramma di interazione del ligando di a QUD, b (2R,3R)-pteroside C, c (3S)-pteroside D, d (2R,3R)-pterosina C, e (2R)-pteroside B, f (2S,3R)-pterosina C, e g (2R)-pterosina B nel sito attivo di BACE1. I colori delle linee tratteggiate spiegano i tipi di varie interazioni: interazioni di legame idrogeno (verde), interazioni idrofobiche (rosa) e interazioni π-sigma (viola). BACE1 β-site amyloid precursor protein cleaving enzyme 1
La posa docked di QUD ha mostrato un’energia di legame (B.E.) di -7.59 kcal/mol. Come mostrato in Fig. 4a, il gruppo NH2 sull’anello chinazolina del ligando ha mostrato due legami a idrogeno con i gruppi CO di Asp32 e Asp228 a distanze di 1,86 e 2,17 Å, rispettivamente. Un terzo legame a idrogeno è stato osservato tra l’altro gruppo NH del ligando e il gruppo CO di Gly230 a una distanza di 2,16 Å. Leu30, Tyr71, Phe108 e Val332 hanno mediato le interazioni idrofobiche. La figura 4b-d mostra le pose docked di (2R,3R)-pteroside C, (3S)-pteroside D e (2R,3R)-pterosina C (inibitori di tipo misto BACE1), rispettivamente. Sono stati posizionati nella tasca di legame rivestita da Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228, e Thr231. Secondo i loro livelli di attività, (2R,3R)-pteroside C (IC50 = 9,74 µM), (3S)-pteroside D (IC50 = 10,7 µM) e (2R,3R)-pterosina C (IC50 = 26,2 µM) hanno mostrato un B.E. di -7,27, -6,93, e -4,84 kcal/mol, rispettivamente. Lo (2R,3R)-Pteroside C ha esibito una potenza maggiore rispetto alla (2R,3R)-pterosina C a causa dell’esistenza di un ulteriore gruppo 2-idrossimetil-tetraidro-pirano-3,4,5-triolo, che ha formato tre legami idrogeno (Fig. 4b). Il gruppo OH di idrossimetile e il gruppo 3-OH dell’anello tetraidro-pirano-triolo hanno mostrato due legami a idrogeno con il gruppo CO di Asp228 a distanze di 2,63 e 2,12 Å, rispettivamente. Inoltre, il gruppo 3-OH ha dimostrato un legame a idrogeno con Thr231 a una distanza di 1,89 Å. Nel caso del (3S)-pteroside D (Fig. 4c), la presenza del gruppo 2,2-dimetilico sull’anello indanonico ha leggermente alterato le interazioni di legame rispetto al (2R,3R)-pteroside C. Il gruppo 3-OH dell’anello tetraidro-piran-triolo ha mostrato solo un singolo legame a idrogeno con Asp228 a una distanza di 1,93 Å. Tuttavia, il gruppo 3-OH dell’anello dell’indanone ha mostrato un ulteriore legame a idrogeno con Ile126 a una distanza di 2,34 Å. Queste interazioni hanno leggermente abbassato l’attività del (3S)-pteroside D rispetto al (2R,3R)-pteroside C. Le interazioni di docking di (2R,3R)-pteroside C visualizzate in Fig. 4d mostrano il gruppo 3-OH dell’anello indanonico legato a Ser36 e Asn37 tramite due legami idrogeno a distanze di 1,83 e 2,07 Å, rispettivamente. Ala39, Val69, Trp76, Ile118, e Arg128 sono stati coinvolti in interazioni idrofobiche, mentre Val69 ha mostrato un’interazione π-sigma.
Le pose agganciate di (2R)-pteroside B, (2S,3R)-pterosin C, e (2R)-pterosin B (inibitori non competitivi di BACE1) sono mostrati in Fig. 4e-g, rispettivamente. Sono stati agganciati nella cavità racchiusa da Asn37, Val69, Tyr71, Trp76, Lys107, Phe108, e Ile126. In accordo con i loro valori di attività, (2R)-pteroside B (IC50 = 18,0 µM), (2S,3R)-pterosina C (IC50 = 23,1 µM), e (2R)-pterosina B (IC50 = 29,6 µM) hanno mostrato un B.E. di -6,16, -5,07, e -4,64 kcal/mol, rispettivamente. Come mostrato in Fig. 4e, il (2R)-pteroside B ha dimostrato una maggiore attività rispetto al (2S,3R)-pteroside C e al (2R)-pteroside B grazie alla presenza di un ulteriore gruppo 2-idrossimetil-tetraidro-pirano-3,4,5-triolo, che ha mostrato quattro interazioni di legame a idrogeno. Due legami a idrogeno sono stati osservati tra il gruppo 4-OH dell’anello tetraidro-triolo e i gruppi NH e CO di Asn37 a distanze di 2,48 e 2,19 Å, rispettivamente. Inoltre, i gruppi 3- e 5-OH hanno mostrato due ulteriori legami a idrogeno con Ile126 e Trp76 a distanze di 2,19 e 2,46 Å, rispettivamente. (2S,3R)-Pterosin C ha mostrato un’attività leggermente migliore di (2R)-pterosin B a causa della presenza di un ulteriore gruppo OH in posizione-3 dell’anello indanone, che ha formato un legame a idrogeno con Lys107 ad una distanza di 2,12 Å (Fig. 4f). Le altre interazioni erano simili a quelle di (2R)-pterosin B. Come mostrato in Fig. 4g, (2R)-pterosin B ha mostrato interazioni idrofobiche con Val69, Tyr76, e Phe108.
AChE docking
(2R)-Pteroside B e (2R,3R)-pterosina C sono stati selezionati come rappresentanti per dimostrare le modalità docked di inibitori AChE di tipo misto e non competitivo, rispettivamente, a causa delle loro attività e tipo di inibizione AChE. Figura 5a, b illustra i modelli di docking di (2R)-pteroside B e (2R,3R)-pterosina C, rispettivamente. Le interazioni dei composti agganciati all’interno del sito attivo di AChE sono mostrate in Fig. 6.

Modelli di docking molecolare per a l’inibitore AChE di tipo misto (2R)-pteroside B (colore blu) e b l’inibitore AChE non competitivo (2R,3R)-pterosina C (colore marrone). Le pose di dock sono sovrapposte alla struttura cristallina a raggi X di E2020 (colore giallo) (codice PDB: 4EY7). AChE, residui del sito attivo e composti sono mostrati da modelli a nastro, linea e bastone, rispettivamente. I colori delle linee tratteggiate spiegano i tipi di varie interazioni: interazioni di legame idrogeno (verde) e interazioni idrofobiche (rosa). AChE acetilcolinesterasi

Diagramma di interazione del ligando di a E2020, b (2R)-pteroside B, c (2R,3R)-pteroside C, d (2S,3R)-pterosina C, e (2R)-pterosina B, f (3S)-pteroside D e g (2R,3R)-pterosina C nel sito attivo di AChE. I colori delle linee tratteggiate spiegano i tipi di varie interazioni: interazioni di legame idrogeno (verde), interazioni idrofobiche (rosa) e interazioni π-sigma (viola). AChE acetilcolinesterasi
La posa docked di E2020 ha dimostrato un B.E. di -10,28 kcal/mol. Come illustrato in Fig. 6a, il gruppo CO dell’anello indanonico ha formato un legame a idrogeno con il gruppo NH di Phe295 a una distanza di 1,70 Å. Trp286 e Tyr341 sono stati coinvolti in interazioni π-sigma, mentre Trp86, Trp286, Tyr337, Phe338, e Tyr341 hanno mediato interazioni idrofobiche. La Figura 6b-f mostra le pose docked di (2R)-pteroside B, (2R,3R)-pteroside C, (2S,3R)-pterosina C, (2R)-pterosina B e (3S)-pteroside D (inibitori AChE di tipo misto), rispettivamente. Sono stati alloggiati nel sito attivo circondato da Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338, e Tyr341. Coerentemente con i loro valori di attività, (2R)-pteroside B (IC50 = 2,55 µM), (2R,3R)-pteroside C (IC50 = 3,77 µM), (2S,3R)-pterosina C (IC50 = 12.8 µM), (2R)-pteroside B (IC50 = 16,2 µM) e (3S)-pteroside D (IC50 = 27,4 µM) hanno mostrato un B.E. di -7,90, -7,49, -6,03, -5,76, e -4,91 kcal/mol, rispettivamente. Lo (2R)-Pteroside B ha dimostrato una potenza maggiore rispetto alla (2S,3R)-pterosina C e alla (2R)-pterosina B grazie alla presenza di un ulteriore gruppo 2-idrossimetil-tetraidro-piran-3,4,5-triolo, che ha stabilito tre interazioni di legame idrogeno (Fig. 6b). Il gruppo OH di idrossimetile e il gruppo 3-OH dell’anello tetraidro-pirano-triolo hanno mostrato legami a idrogeno con il gruppo CO di Asn87 a distanze di 2,23 e 2,28 Å, rispettivamente. Inoltre, il gruppo 4-OH ha formato un legame a idrogeno con il gruppo CO di Trp86 a una distanza di 2,13 Å. Nel caso di (2R,3R)-pteroside C (Fig. 6c), il gruppo metilico in posizione-2 dell’anello indanone non ha mostrato un’interazione idrofobica con Tyr341 e quindi ha mostrato un’attività relativamente più bassa rispetto al (2R)-pteroside B. Tuttavia, Tyr341 ha mantenuto l’interazione idrofobica con l’altra parte dell’anello indanone come mostrato in quello di (2R)-pteroside B. La maggiore attività di (2S,3R)-pterosina C rispetto a (2R)-pterosina B è stata attribuita all’esistenza di un ulteriore gruppo OH in posizione-3 dell’anello indanone, che ha formato un legame idrogeno con Phe295 ad una distanza di 1,82 Å (Fig. 6d). Le interazioni rimanenti erano paragonabili alle interazioni (2R)-pterosina B. Come mostrato in Fig. 6e, il gruppo OH del gruppo idrossietilico in posizione-6 dell’anello indanone formato un legame a idrogeno con Tyr124 ad una distanza di 2,48 Å. Tyr124, Trp286, Tyr337, Phe338, e Tyr341 contribuito alle interazioni idrofobiche. Rispetto a (2R)-pteroside B e (2R,3R)-pteroside C, (3S)-pteroside D ha mostrato interazioni di legame dissimili a causa della presenza del gruppo 2,2-dimetilico all’anello indanone (Fig. 6f). Il gruppo 2,2-dimetilico ha contribuito significativamente alla distinta posizione docked del (3S)-pteroside D. Il gruppo 5-OH dell’anello tetraidro-piran-triolo ha formato un legame idrogeno con Asp74 ad una distanza di 1,96 Å. Il gruppo CO dell’anello dell’indanone ha mostrato un legame a idrogeno con Tyr72 a una distanza di 2,97 Å. Queste interazioni spiegano la bassa attività del (3S)-pteroside D.
La posizione docked della (2R,3R)-pterosina C (inibitore AChE non competitivo) è mostrata in Fig. 6g. (2R,3R)-Pterosina C (IC50 = 23,2 µM) ha dimostrato un B.E. di -5,01 kcal/mol. La tasca di legame della (2R,3R)-pterosina C comprendeva Tyr72, Trp286, Ser293, Phe297 e Tyr341, con due interazioni di legame a idrogeno. Uno dei legami a idrogeno si è formato tra il gruppo 3-OH dell’anello indanonico e il gruppo CO di Ser293 a una distanza di 2,03 Å. Il secondo legame a idrogeno è stato osservato tra il gruppo OH del gruppo idrossietilico presente in posizione 6 dell’anello indanonico e Tyr72 a una distanza di 1,94 Å. Residui come Tyr72, Trp286, Phe297, e Tyr341 hanno partecipato alle interazioni idrofobiche.
BChE docking
Considerando i livelli di attività e il tipo di inibizione di BChE, (2R,3R)-pteroside C e (2S,3R)-pterosin C sono stati selezionati per dimostrare le modalità docked degli inibitori BChE di tipo misto e non competitivo, rispettivamente. La figura 7a, b illustra i modelli di docking di (2R,3R)-pteroside C e (2S,3R)-pterosina C, rispettivamente. Le interazioni dei composti agganciati all’interno del sito attivo di BChE sono presentate in Fig. 8.
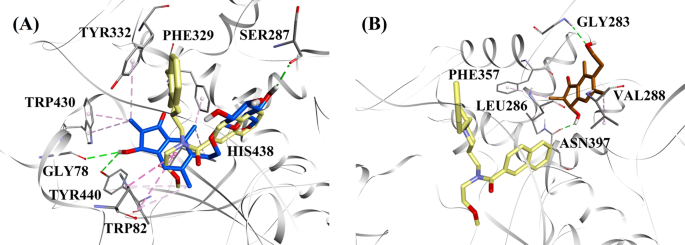
Modelli di docking molecolare per a l’inibitore BChE di tipo misto (2R,3R)-pteroside C (colore blu) e b l’inibitore BChE non competitivo (2S,3R)-pterosina C (colore marrone). Le pose di dock sono sovrapposte alla struttura cristallina a raggi X di 3F9 (colore giallo) (codice PDB: 4TPK). BChE, residui del sito attivo e composti sono mostrati rispettivamente da modelli a nastro, linea e bastone. I colori delle linee tratteggiate spiegano i tipi di varie interazioni: interazioni di legame a idrogeno (verde) e interazioni idrofobiche (rosa). BChE butilcolinesterasi
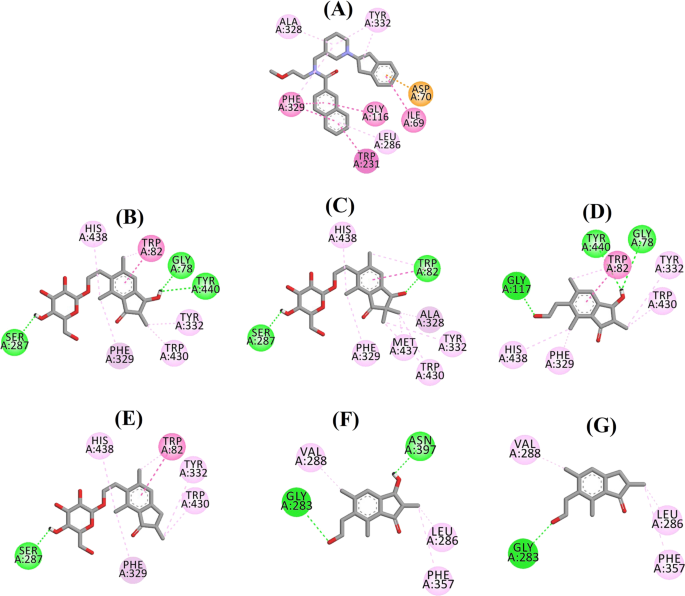
Diagramma di interazione del ligando di a 3F9, b (2R,3R)-pteroside C, c (3S)-pteroside D, d (2R,3R)-pterosina C, e (2R)-pteroside B, f (2S,3R)-pterosina C e g (2R)-pterosina B nel sito attivo di BChE. I colori delle linee tratteggiate spiegano i tipi di varie interazioni: interazioni di legame idrogeno (verde), interazioni idrofobiche (rosa) e interazioni π-anione (oro). BChE butilcolinesterasi
La posa docked della 3F9 ha mostrato un B.E. di -8,49 kcal/mol. Come mostrato in Fig. 8a, le interazioni idrofobiche erano principalmente responsabili del legame del ligando. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329, e Tyr332 rappresentato per le interazioni idrofobiche, mentre Asp70 dimostrato una interazione π-anione. Figura 8b-e illustra le pose docked di (2R,3R)-pteroside C, (3S)-pteroside D, (2R,3R)-pterosina C e (2R)-pteroside B (inibitori BChE di tipo misto), rispettivamente. La loro tasca di legame era composta da Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438, e Tyr440. In accordo con i loro livelli di attività, (2R,3R)-pteroside C (IC50 = 5,29 µM), (3S)-pteroside D (IC50 = 19,3 µM), (2R,3R)-pterosina C (IC50 = 20,3 µM) e (2R)-pteroside B (IC50 = 62,0 µM) hanno dimostrato un B.E. di -7,23, -6,59, -6,52, e -4,38 kcal/mol, rispettivamente. Come mostrato in Fig. 8b, il gruppo 3-OH dell’anello indanonico del (2R,3R)-pteroside C ha mostrato due legami idrogeno con Gly78 e Tyr440 a distanze di 2,87 e 2,89 Å, rispettivamente. Nel caso del (3S)-pteroside D, il gruppo 2,2-dimetilico all’anello indanonico ha influenzato le interazioni di legame del gruppo 3-OH (Fig. 8c). Il gruppo 3-OH ha formato solo un legame a idrogeno con Trp82 a una distanza di 2,94 Å, che ha portato ad una attività relativamente più bassa di (3S)-pteroside D rispetto a (2R,3R)-pteroside C. Come mostrato in Fig. 8d, (2R,3R)-pterosin C non è riuscito a produrre un legame a idrogeno con Ser287 a causa dell’assenza di un gruppo 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol. Di conseguenza, ha prodotto un’attività inferiore rispetto al (2R,3R)-pteroside C. La mancanza del gruppo OH in posizione-3 dell’anello indanonico è risultata essere responsabile dell’attività molto bassa del (2R)-pteroside B (Fig. 8e), che non è riuscito a formare legami a idrogeno con Gly78 e Tyr440 come il (2R,3R)-pteroside C.
Le posizioni agganciate di (2S,3R)-pterosina C e (2R)-pterosina B (inibitori non competitivi di BChE) sono mostrate in Fig. 8f, g, rispettivamente. Queste pose agganciate erano contenute nella cavità racchiusa da Gly283, Leu286, Val288, Phe357, e Asn397. Per quanto riguarda i loro livelli di attività, la (2S,3R)-pterosina C (IC50 = 44,3 µM) e la (2R)-pterosina B (IC50 = 48,1 µM) hanno mostrato un B.E. di -5,40 e -5,06 kcal/mol, rispettivamente. La (2S,3 R)-Pterosina C era più potente della (2R)-pterosina B a causa della presenza di un ulteriore gruppo OH in posizione-3 dell’anello indanonico, che formava un legame a idrogeno con il gruppo CO di Asn397 a una distanza di 2,05 Å (Fig. 8f). Altre interazioni sono state trovate simili a (2R)-pterosina B. Come mostrato in Fig. 8g, il gruppo OH nel gruppo idrossietilico in posizione-6 dell’anello indanonico ha formato un legame a idrogeno con Gly283 a una distanza di 2,40 Å. I residui Leu286, Val288, e Phe357 hanno partecipato alle interazioni idrofobiche.
Gli inibitori di tipo misto si legano sia all’enzima libero che al complesso enzima-substrato, il che indica che questi composti possono legarsi al sito catalitico di ciascun enzima corrispondente. Gli inibitori non competitivi si legano al sito allosterico dell’enzima libero o del complesso enzima-substrato. Uno studio recente ha suggerito che gli inibitori competitivi, di tipo misto e non competitivi occupano siti diversi nelle tasche di legame di BACE1, AChE e BChE38. Durante il docking per la valutazione del meccanismo inibitorio dei derivati della pterosina, i siti di legame dei composti sono stati definiti secondo il loro tipo di inibizione. I risultati del docking hanno indicato che i siti di legame degli inibitori di tipo misto e non competitivo per BACE1, AChE e BChE si sovrappongono parzialmente in ogni sito attivo corrispondente e sono coerenti con un rapporto precedente38.
Permeabilità della BBB
PAMPA-BBB, un test di permeabilità di membrana artificiale in vitro per la BBB, è uno degli strumenti di screening fisico-chimico più affidabili nella scoperta iniziale di farmaci mirati al SNC40. Il sistema PAMPA-BBB modella la diffusione passiva transcellulare delle sostanze chimiche attraverso la BBB e misura i meccanismi di trasporto strettamente passivi attraverso una membrana lipidica artificiale sulla permeabilità effettiva (Pe, cm/s). Sulla base del modello stabilito per la previsione della permeazione della BBB, i composti sono stati classificati in (i) “CNS+” (alta permeazione della BBB prevista); Pe (10-6 cm/s) > 4.00, (ii) “CNS-” (bassa permeazione della BBB prevista); Pe (10-6 cm/s) < 2.00, e (iii) “CNS+/-” (permeazione della BBB incerta); Pe (10-6 cm/s) da 4.00 a 2.00. Di conseguenza, (2R)-pterosina B, (2S)-pterosina P, e (2S)-pterosina A esposto alta permeazione BBB con valori Pe di 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s, e 6,26 × 10-6 cm/s, rispettivamente (Tabella 4, Informazioni supplementari 4). Il valore Pe di (2R)-pterosin B era 1,7 volte superiore a quello del verapamil farmaco CNS (Pe = 34,6 × 10-6 cm/s), che è stato utilizzato per il controllo positivo. (2S,3R)-Pterosina C e (2R,3R)-pterosina C hanno mostrato una permeazione incerta della BBB con valori Pe di 2,34 e 1,98, rispettivamente. (2R,3R)-Pteroside C, (3S)-pteroside D, e (2R)-pteroside B, che hanno mostrato le attività inibitorie di BACE1 e colinesterasi più potenti tra i derivati della pterosina testati, hanno mostrato una permeabilità BBB molto bassa. L’esistenza del gruppo 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol come nei derivati della pteroside ha portato a una notevole diminuzione della permeabilità della BBB. Rispetto alla (2R)-pterosina B, la presenza aggiuntiva del gruppo OH in posizione-3 dell’anello indanonico come nella pterosina C, il gruppo idrossimetilico in posizione-2 dell’anello indanonico come nella (2S)-pterosina A o il gruppo idrossimetilico in posizione-5 dell’anello indanonico come nella (2S)-pterosina P ha anche ridotto significativamente la permeabilità della BBB. Considerando una permeabilità della BBB eccezionalmente alta e l’inibizione significativa di BACE1, AChE e BChE, la (2R)-pterosina B può avere il potenziale per esibire una forte attività anti-AD.
Effetti della (2R)-pterosina B e (2R,3R)-pteroside C sulla secrezione di peptidi Aβ da parte delle cellule neuronali
Per studiare la funzione della (2R)-pterosina B e (2R,3R)-pteroside C nel diminuire l’escrezione di Aβ dalle cellule neuronali, abbiamo usato una linea cellulare di neuroblastoma murino che sovraesprime in modo stabile APPswe umana. La linea cellulare è un modello cellulare di AD caratterizzato dalla secrezione eccessiva di Aβ40 e Aβ42. Gli oligomeri amiloidi tossici sono formati dalle due isoforme del peptide Aβ con lunghezze diverse. Aβ40 è l’isoforma Aβ più abbondante nel cervello41, mentre Aβ42 aumenta significativamente con alcune forme di AD42. Sandwich ELISA di Aβ40 ha mostrato che (2R)-pterosin B significativamente ridotto la quantità di peptide Aβ40 secreto dalle cellule di neuroblastoma in media fino al 50% a 500 μM (P < 0,01) (Fig. 9a). Allo stesso modo, la secrezione del peptide Aβ42 da parte delle cellule di neuroblastoma è diminuita significativamente in presenza di 500 µM di (2R,3R)-pteroside C (P < 0,05) (Fig. 9b). In conclusione, la (2R)-pterosina B e la (2R,3R)-pteroside C hanno diminuito significativamente la secrezione di peptidi Aβ dalle cellule di neuroblastoma ad una concentrazione di 500 μM.

a Effetto della (2R)-pterosina B sulla secrezione di peptidi Aβ. Le cellule di neuroblastoma secernenti APPswe sono state trattate con 12, 60, 250 o 500 µM di (2R)-pterosina B per 24 ore e i mezzi condizionati sono stati raccolti in presenza di un inibitore della proteasi. β-SI (10 µM) è stato usato come controllo positivo. Il controllo negativo includeva cellule coltivate in assenza di composti di prova. L’analisi quantitativa di Aβ40 e Aβ42 secreti nei mezzi condizionati è stata eseguita usando un ELISA a sandwich. Le medie ± SD da tre esperimenti indipendenti sono mostrati. I peptidi Aβ40 secreti sono diminuiti significativamente in presenza di 500 µM (2R)-pterosina B. CON controllo negativo, β-SI β-secretasi inibitore III, Aβ β-amiloide. *P < 0,05, **P < 0,01. b Effetto di (2R,3R)-pteroside C sulla secrezione di peptidi Aβ. Le cellule di neuroblastoma secernenti APPswe sono state trattate con 12, 60, 125, 250 o 500 µM di (2R,3R)-pteroside C per 24 ore, e i mezzi condizionati sono stati raccolti in presenza di un inibitore della proteasi. Il β-SI (10 µM) è stato usato come controllo positivo. Il controllo negativo includeva cellule coltivate in assenza di composti di prova. L’analisi quantitativa di Aβ40 e Aβ42 secreto nei mezzi condizionati è stata eseguita utilizzando sandwich ELISA. Le medie ± SD da tre esperimenti indipendenti sono mostrati. Il peptide Aβ42 secreto è diminuito significativamente in presenza di 500 µM (2R,3R)-pteroside C. CON controllo negativo, β-SI β-secretasi inibitore III, Aβ β-amiloide. *P < 0.05
Citotossicità dei derivati della pterosina basata sul test MTT43
In generale, i derivati della pterosina hanno mostrato una citotossicità trascurabile contro varie linee cellulari normali e tumorali, come il neuroblastoma umano SH-SY5Y, le cellule gliali di ratto C6, i fibroblasti embrionali di topo NIH3T3 e il melanoma di topo B16F10 con valori LD50 sopra 0.5 mM (Informazioni supplementari 5). In particolare, (2R)-pterosina B, (3R)-pterosina D, (2S)-pterosina P, (2S)-pteroside A, (2R)-pteroside B, e (2R,3R)-pteroside C non ha mostrato citotossicità contro le linee cellulari testate con valori LD50 sopra 5 mM. Curiosamente, diverse pterosine hanno mostrato effetti antiproliferativi relativi contro le cellule neuronali SH-SY5Y rispetto alle cellule gliali C6 di origine mesenchimale. I presenti risultati sono stati coerenti con un rapporto precedente che ha suggerito i derivati della pterosina non sono tossici per gli esseri umani26.
.