
Isolatie en karakterisering van pterosinederivaten uit P. aquilinum
Pterosinederivaten werden geïsoleerd uit de gehele planten van P. aquilinum. Voor het onderzoek van de fytochemische bestanddelen van de varen werd een waterextract achtereenvolgens gepartitioneerd met ethylacetaat (EtOAc) en n-butanol (n-BuOH) (Fig. 1). Herhaalde kolomchromatografie van de EtOAc-oplosbare fractie resulteerde in de isolatie van negen derivaten, waaronder (2R)-pterosine B (1), pterosine Z (2), (2S)-pterosine P (3), (3R)-pterosine D (4), (2S)-pterosine A (5), (2S,3R)-pterosine C (6), (2R,3R)-pterosine C (7), (2R)-pteroside B (8), en pteroside Z (9), met zuiverheden van meer dan 97% (Fig. 2). De herhaalde kolomchromatografische scheiding van de n-BuOH oplosbare fractie resulteerde in de isolatie van zes derivaten, waaronder (2S)-pteroside A (10), (2S)-pteroside A2 (11), (2S,3R)-pteroside C (12), (2R,3R)-pteroside C (13), (3S)-pteroside D (14), en (2S)-pteroside P (15), met zuiverheden groter dan 97%. De structuren van de verbindingen werden geïdentificeerd door de analyse van spectrale gegevens, waaronder MS, 1D- en 2D-NMR (Supplementary Information 1).

Structuren van pterosinverbindingen 1-15xxx
Remmende activiteit van pterosinederivaten tegen BACE1, AChE, en BChE
Om het anti-AD potentieel te evalueren, werd de remmende activiteit van elke pterosinverbinding tegen BACE1 en cholinesterases geëvalueerd door middel van respectievelijke in vitro remmingstests (tabel 1). Alle geteste pterosinederivaten vertoonden concentratie-afhankelijke remmende activiteiten tegen BACE1 met een bereik van IC50 waarden (half-maximum remmende concentratie) van 9,74-94,4 μM, met uitzondering van (2S)-pterosine A en (2S)-pteroside P die inactief waren bij de geteste concentraties, vergeleken met de IC50 van quercetine gebruikt als de positieve controle, die 18,8 μM bedroeg. De remmende potentie van de sterkste remmers was in de volgorde van (2R,3R)-pteroside C, (3S)-pteroside D, (2R)-pteroside B, (2S,3R)-pterosine C, (2R,3R)-pterosine C, (2S,3R)-pteroside C, en (2R)-pterosine B met IC50 waarden van 9.74, 10,7, 18,0, 23,1, 26,2, 28,9, en 29,6 μM, respectievelijk. Vervolgens testten we het remmend vermogen van de pterosinederivaten tegen AChE. Alle geteste verbindingen vertoonden significante AChE-remmende activiteiten, met IC50 waarden in het bereik van 2,55-110 μM, vergeleken met de IC50 tegen AChE van berberine, gebruikt als positieve controle, die 0,39 μM was. De pterosine verbindingen die de sterkste remmende activiteit tegen AChE vertoonden waren (2R)-pteroside B, (2R,3R)-pteroside C, (2S,3R)-pteroside C, (2S,3R)-pterosine C, en (2R)-pterosine B met IC50 waarden van respectievelijk 2,55, 3,77, 9,17, 12,8, en 16,2 μM. Tenslotte testten we de remmende capaciteiten van de pterosinederivaten tegen BChE. Alle geteste pterosine verbindingen vertoonden remmende activiteit tegen BChE, met IC50 waarden die varieerden van 5,29 tot 119 μM, met uitzondering van (3R)-pterosine D die inactief was bij de geteste concentraties, vergeleken met de IC50 van berberine tegen BChE, die 3,32 μM bedroeg. De pterosinverbindingen die de sterkste remmende activiteit tegen BChE vertoonden waren (2R,3R)-pteroside C en pteroside Z met IC50-waarden van respectievelijk 5,29 en 5,31 μM.
Collectief vertoonden de meeste geteste pterosinederivaten significante remmende activiteiten tegen BACE1, AChE en BChE tegelijk. De aanwezigheid van de extra 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolgroep zoals in pterosidederivaten verhoogde de remmende activiteiten tegen de enzymen aanzienlijk. Bovendien verminderde de aanwezigheid van de extra hydroxymethylgroep op positie 2 van de indanonring van (2R)-pterosine B zoals in (2S)-pterosine A of de methylgroep zoals in (3R)-pterosine D en pterosine Z de remmende activiteiten tegen de enzymen. Met name de aanwezigheid van de hydroxymethylgroep op positie-5 van de indanonring zoals in (2S)-pterosine P verminderde de remmende activiteit tegen BACE1.
Kinetische parameters van enzymremming door pterosinederivaten
In een poging de wijze van enzymatische remming van pterosinederivaten te verklaren, voerden wij een kinetische analyse van BACE1 en cholinesterases uit voor representatieve remmers (tabel 2, aanvullende informatie 2). Een lage Ki (remmingsconstante) wijst op een nauwere binding van het enzym en een meer doeltreffende remmer. Over het algemeen correleerden de Ki waarden van de verbindingen met de respectievelijke IC50 waarden. De remming van BACE1 door de verbindingen (2R,3R)-pteroside C, (3S)-pteroside D, en (2R,3R)-pterosine C was van het gemengde type met Ki waarden van respectievelijk 12,6, 16,5, en 27.6 µM, respectievelijk, terwijl remming door (2R)-pteroside B, (2S,3R)-pterosine C, en (2R)-pterosine B niet-concurrerend was met Ki waarden van respectievelijk 23,1, 33,8, en 38,3 µM. AChE-remming door (2R)-pteroside B, (2R,3R)-pterosine C, (2S,3R)-pterosine C, en (3S)-pteroside D was van het gemengde type met Ki waarden van 4.89, 8.13, 12.1, 16.3, en 23.1 µM, respectievelijk, terwijl (2R,3R)-pterosine C een niet-competitieve type remmer was met een Ki waarde van 29.6 µM. BChE remming door (2R,3R)-pterosine C, (2R,3R)-pteroside C, (3S)-pteroside D, en (2R)-pteroside B was van het gemengde type met Ki waarden van 4.77, 9.62, 19.7, en 22.6 µM, respectievelijk, terwijl (2S,3R)-pterosine C en (2R)-pterosine B niet-competitieve remmers waren met Ki waarden van 29.9 en 53.5 µM, respectievelijk. Deze resultaten suggereerden dat specifieke pterosinederivaten effectieve BACE1-, AChE- en BChE-remmers zouden kunnen zijn.
Moleculaire docking-simulaties voor BACE1, AChE en BChE
Verschillende kristalstructuren zijn beschikbaar voor BACE1 en cholinesterasen. We selecteerden menselijke PDB’s op basis van wild-type structuren, cocrystallized liganden en resoluties van de structuren. X-straal kristalstructuren van BACE1 gecomplexeerd met QUD (PDB code: 2WJO, resolutie: 2.5 Å)33, AChE gecomplexeerd met E2020 (PDB code: 4EY7, resolutie: 2.35 Å)34, en BChE gecomplexeerd met 3F9 (PDB code: 4TPK, resolutie: 2.70 Å)35 werden geselecteerd voor docking. In eerste instantie werden QUD, E2020 en 3F9 uit de kristalstructuren geëxtraheerd en in de actieve sites van respectievelijk BACE1, AChE en BChE gedockt. Vervolgens werden (2R,3R)-pteroside C, (3S)-pteroside D, (2R,3R)-pterosine C, (2R)-pteroside B, (2S,3R)-pterosine C en (2R)-pterosine B met het bekende remmingsmechanisme tegen BACE1, AChE en BChE gedockt om hun SAR te bepalen. De docking resultaten zijn samengevat in Tabel 3. De SAR van de geselecteerde mixed-type en niet-competitieve BACE1, AChE en BChE remmers maakte de evaluatie mogelijk van nieuwe bindingsplaatsgerichte remming van de enzymen door pterosinen.
Onze dockingmodus van E2020 was consistent met de experimenteel bepaalde bindingsmodus die eerder was gerapporteerd met recombinant menselijk AChE (rhAChE) (Supplementary Information 3)34. De RMSD (root-mean-square-deviatie) tussen het kristal en de gedockte conformaties van E2020 was 0,54 Å, wat suggereert dat onze docking-opstelling betrouwbaar is voor het reproduceren van de experimentele bindingsmodus. Bovendien leidde de docked mode van E2020 tot een vergelijkbare interactie als die van rhAChE-E2020. In onze studie werden watermoleculen uit de kristalstructuur verwijderd tijdens het dokken; daarom werden water-gemedieerde interacties niet geanalyseerd in de huidige studie. Evenzo waren de gedockte modi van QUD en 3F9 consistent met de beschikbare experimentele gegevens voor respectievelijk BACE1 33 en BChE35 (Supplementary Information 3). De RMSD’s tussen de kristalconformatie en de gedockte conformatie van QUD en 3F9 waren respectievelijk 0,46 en 0,60 Å. Verder waren de bindingsplaatsen van pterosineremmers in overeenstemming met een eerdere dockingstudie waarbij BACE1, AChE en BChE38 betrokken waren. In deze studie werd echter gebruik gemaakt van Tetronarce californica AChE (PDB-code: 1ACJ), dat iets andere residunummers bevat dan menselijk AChE als gevolg van variaties in hun sequenties.
BACE1-docking
Gebaseerd op het type remming en de activiteit, werden (2R,3R)-pteroside C en (2R)-pteroside B geselecteerd om de gedockte modi van respectievelijk mixed-type en noncompetitieve BACE1-remmers aan te tonen. Figuur 3a, b toont de docking-modellen van respectievelijk (2R,3R)-pteroside C en (2R)-pteroside B. De interacties van de gedockte verbindingen in de actieve site van BACE1 worden getoond in Fig. 4.

Moleculaire dockingmodellen voor a de BACE1-remmer van het gemengde type (2R,3R)-pteroside C (blauwe kleur) en b de niet-concurrerende BACE1-remmer (2R)-pteroside B (bruine kleur). De gedockte poses zijn gesuperponeerd op de X-straal kristalstructuur van QUD (gele kleur) (PDB-code: 2WJO). BACE1, actieve site residuen en verbindingen worden getoond door respectievelijk lint, lijn en stick modellen. De kleuren van de stippellijnen verklaren de types van verschillende interacties: waterstofbruginteracties (groen), hydrofobe interacties (roze) en π-sigma interacties (paars). BACE1 β-site amyloïd precursor proteïne klievend enzym 1

Ligand-interactiediagram van a QUD, b (2R,3R)-pteroside C, c (3S)-pteroside D, d (2R,3R)-pterosine C, e (2R)-pteroside B, f (2S,3R)-pterosine C, en g (2R)-pterosine B in de actieve site van BACE1. De kleuren van de stippellijnen verklaren de types van de verschillende interacties: waterstofbruginteracties (groen), hydrofobe interacties (roze) en π-sigma interacties (paars). BACE1 β-site amyloïd precursor proteïne klievend enzym 1
De gedockte pose van QUD vertoonde een bindingsenergie (B.E.) van -7,59 kcal/mol. Zoals te zien is in Fig. 4a, vertoonde de NH2 groep op de quinazoline ring van het ligand twee waterstofbruggen met de CO groepen van Asp32 en Asp228 op afstanden van respectievelijk 1.86 en 2.17 Å. Een derde waterstofbrug werd waargenomen tussen de andere NH groep van het ligand en de CO groep van Gly230 op een afstand van 2.16 Å. Leu30, Tyr71, Phe108, en Val332 bemiddelden de hydrofobe interacties. Figuur 4b-d toont de gedokte poses van respectievelijk (2R,3R)-pteroside C, (3S)-pteroside D en (2R,3R)-pterosine C (remmers van het gemengde type BACE1). Zij werden gepositioneerd in de bindingszak omgeven door Ser36, Asn37, Ala39, Val69, Tyr71, Trp76, Ile118, Ile126, Arg128, Asp228, en Thr231. Wat hun activiteitsniveaus betreft, vertoonden (2R,3R)-pteroside C (IC50 = 9,74 µM), (3S)-pteroside D (IC50 = 10,7 µM) en (2R,3R)-pterosine C (IC50 = 26,2 µM) een B.E. van respectievelijk -7,27, -6,93, en -4,84 kcal/mol. (2R,3R)-Pteroside C vertoonde een hogere potentie dan (2R,3R)-pterosine C door de aanwezigheid van een extra 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolgroep, die drie waterstofbruggen vormde (Fig. 4b). De OH-groep van hydroxymethyl en de 3-OH-groep van de tetrahydro-pyran-triol ring vertoonden twee waterstofbruggen met de CO-groep van Asp228 op afstanden van respectievelijk 2,63 en 2,12 Å. Bovendien vertoonde de 3-OH groep een waterstofbrug met Thr231 op een afstand van 1,89 Å. In het geval van (3S)-pteroside D (Fig. 4c), veranderde de aanwezigheid van de 2,2-dimethylgroep aan de indanonring enigszins de bindingsinteracties vergeleken met (2R,3R)-pteroside C. De 3-OH groep van de tetrahydro-pyran-triol ring vertoonde slechts een enkele waterstofbrug met Asp228 op een afstand van 1,93 Å. De 3-OH-groep van de indanonring vertoonde echter een extra waterstofbrug met Ile126 op een afstand van 2,34 Å. Deze interacties verlaagden de activiteit van (3S)-pteroside D enigszins ten opzichte van (2R,3R)-pteroside C. De docking interacties van (2R,3R)-pterosine C weergegeven in Fig. 4d tonen de 3-OH groep van de indanon ring gebonden aan Ser36 en Asn37 via twee waterstofbruggen op afstanden van respectievelijk 1.83 en 2.07 Å. Ala39, Val69, Trp76, Ile118, en Arg128 waren betrokken bij hydrofobe interacties, terwijl Val69 een π-sigma interactie vertoonde.
De gedokte poses van (2R)-pteroside B, (2S,3R)-pterosine C, en (2R)-pterosine B (niet-competitieve BACE1 remmers) worden getoond in Fig. 4e-g, respectievelijk. Zij werden vastgezet in de holte omsloten door Asn37, Val69, Tyr71, Trp76, Lys107, Phe108, en Ile126. In overeenstemming met hun activiteitswaarden vertoonden (2R)-pteroside B (IC50 = 18,0 µM), (2S,3R)-pterosine C (IC50 = 23,1 µM), en (2R)-pterosine B (IC50 = 29,6 µM) een B.E. van respectievelijk -6,16, -5,07, en -4,64 kcal/mol. Zoals weergegeven in Fig. 4e, vertoonde (2R)-pteroside B een hogere activiteit dan (2S,3R)-pterosine C en (2R)-pterosine B door de aanwezigheid van een extra 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol groep, die vier waterstofbrug interacties vertoonde. Twee waterstofbruggen werden waargenomen tussen de 4-OH groep van de tetrahydro-pyran-triol ring en de NH en CO groepen van Asn37 op afstanden van respectievelijk 2,48 en 2,19 Å. Verder vertoonden de 3- en 5-OH groepen twee extra waterstofbruggen met Ile126 en Trp76 op afstanden van respectievelijk 2.19 en 2.46 Å. (2S,3R)-Pterosine C vertoonde een iets betere activiteit dan (2R)-pterosine B door de aanwezigheid van een extra OH-groep op positie-3 van de indanonring, die een waterstofbrug vormde met Lys107 op een afstand van 2,12 Å (Fig. 4f). De andere interacties waren vergelijkbaar met die van (2R)-pterosine B. Zoals weergegeven in Fig. 4g, vertoonde (2R)-pterosine B hydrofobe interacties met Val69, Tyr76, en Phe108.
AChE-docking
(2R)-Pteroside B en (2R,3R)-pterosine C werden geselecteerd als vertegenwoordigers om de gedockte modi van respectievelijk mixed-type en niet-competitieve AChE-remmers aan te tonen, vanwege hun activiteiten en type van AChE-remming. Figuur 5a, b toont de docking-modellen van respectievelijk (2R)-pteroside B en (2R,3R)-pterosine C. De interacties van de verbindingen in de actieve site van AChE worden getoond in Fig. 6.

Moleculaire dockingmodellen voor a de mixed-type AChE-remmer (2R)-pteroside B (blauwe kleur) en b de niet-competitieve AChE-remmer (2R,3R)-pterosine C (bruine kleur). De gedockte poses zijn gesuperponeerd op de X-straal kristalstructuur van E2020 (gele kleur) (PDB-code: 4EY7). AChE, actieve site residuen en verbindingen worden getoond door respectievelijk lint-, lijn- en stick-modellen. De kleuren van de stippellijnen verklaren het type van de verschillende interacties: waterstofbruginteracties (groen) en hydrofobe interacties (roze). AChE acetylcholinesterase

Ligandinteractiediagram van a E2020, b (2R)-pteroside B, c (2R,3R)-pteroside C, d (2S,3R)-pterosine C, e (2R)-pterosine B, f (3S)-pteroside D en g (2R,3R)-pterosine C in de actieve site van AChE. De kleuren van de stippellijnen verklaren het type van de verschillende interacties: waterstofbruginteracties (groen), hydrofobe interacties (roze) en π-sigma interacties (paars). AChE acetylcholinesterase
De gedockte pose van E2020 vertoonde een B.E. van -10,28 kcal/mol. Zoals geïllustreerd in Fig. 6a, vormde de CO groep van de indanon ring een waterstofbrug met de NH groep van Phe295 op een afstand van 1.70 Å. Trp286 en Tyr341 waren betrokken bij π-sigma interacties, terwijl Trp86, Trp286, Tyr337, Phe338, en Tyr341 hydrofobe interacties bemiddelden. Figuur 6b-f toont de gedokte poses van respectievelijk (2R)-pteroside B, (2R,3R)-pteroside C, (2S,3R)-pterosine C, (2R)-pterosine B en (3S)-pteroside D (mixed-type AChE remmers). Zij werden ondergebracht in de actieve site omgeven door Tyr72, Asp74, Trp86, Asn87, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338, en Tyr341. In overeenstemming met hun activiteitswaarden werden (2R)-pteroside B (IC50 = 2,55 µM), (2R,3R)-pteroside C (IC50 = 3,77 µM), (2S,3R)-pterosine C (IC50 = 12.8 µM), (2R)-pterosine B (IC50 = 16,2 µM) en (3S)-pteroside D (IC50 = 27,4 µM) vertoonden een B.E. van respectievelijk -7,90, -7,49, -6,03, -5,76, en -4,91 kcal/mol. (2R)-Pteroside B vertoonde een hogere potentie dan (2S,3R)-pterosine C en (2R)-pterosine B door de aanwezigheid van een extra 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triol groep, die drie waterstofbrug interacties tot stand bracht (Fig. 6b). De OH-groep van hydroxymethyl en de 3-OH-groep van de tetrahydro-pyran-triol ring vertoonden waterstofbruggen met de CO-groep van Asn87 op afstanden van respectievelijk 2,23 en 2,28 Å. Verder vormde de 4-OH groep een waterstofbrug met de CO groep van Trp86 op een afstand van 2.13 Å. In het geval van (2R,3R)-pteroside C (Fig. 6c), vertoonde de methylgroep op positie-2 van de indanonring geen hydrofobe interactie met Tyr341 en vertoonde dus een relatief lagere activiteit dan (2R)-pteroside B. Tyr341 handhaafde echter de hydrofobe interactie met het andere deel van de indanonring, zoals aangetoond in die van (2R)-pteroside B. De hogere activiteit van (2S,3R)-pterosine C dan (2R)-pterosine B werd toegeschreven aan het bestaan van een extra OH-groep op positie-3 van de indanonring, die een waterstofbrug vormde met Phe295 op een afstand van 1,82 Å (Fig. 6d). De overige interacties waren vergelijkbaar met (2R)-pterosine B interacties. Zoals te zien in Fig. 6e, vormde de OH groep van de hydroxyethyl groep op positie-6 van de indanon ring een waterstofbrug met Tyr124 op een afstand van 2.48 Å. Tyr124, Trp286, Tyr337, Phe338, en Tyr341 droegen bij aan de hydrofobe interacties. Vergeleken met (2R)-pteroside B en (2R,3R)-pteroside C, vertoonde (3S)-pteroside D ongelijksoortige bindingsinteracties door de aanwezigheid van de 2,2-dimethylgroep aan de indanonring (Fig. 6f). De 2,2-dimethylgroep droeg significant bij aan de aparte docked pose van (3S)-pteroside D. De 5-OH groep van de tetrahydro-pyran-triol ring vormde een waterstofbrug met Asp74 op een afstand van 1,96 Å. De CO groep van de indanon ring vertoonde een waterstofbrug met Tyr72 op een afstand van 2.97 Å. Deze interacties verklaarden de lage activiteit van (3S)-pteroside D.
De gedockte pose van (2R,3R)-pterosine C (niet-competitieve AChE remmer) wordt getoond in Fig. 6g. (2R,3R)-Pterosine C (IC50 = 23,2 µM) vertoonde een B.E. van -5,01 kcal/mol. De bindingszak van (2R,3R)-pterosine C bestond uit Tyr72, Trp286, Ser293, Phe297, en Tyr341, met twee waterstofbruginteracties. Eén van de waterstofbruggen werd gevormd tussen de 3-OH groep van de indanonring en de CO groep van Ser293 op een afstand van 2.03 Å. De tweede waterstofbrug werd waargenomen tussen de OH groep van de hydroxyethylgroep aanwezig op positie-6 van de indanonring en Tyr72 op een afstand van 1.94 Å. Residuen zoals Tyr72, Trp286, Phe297, en Tyr341 namen deel aan hydrofobe interacties.
BChE docking
Gezien de activiteitsniveaus en het type BChE remming, werden (2R,3R)-pteroside C en (2S,3R)-pterosine C geselecteerd om de gedokte modi van respectievelijk mixed-type en niet-competitieve BChE remmers aan te tonen. Figuur 7a, b toont de docking-modellen van respectievelijk (2R,3R)-pteroside C en (2S,3R)-pterosine C. De interacties van de gedockte verbindingen binnen de actieve site van BChE worden gepresenteerd in Fig. 8.
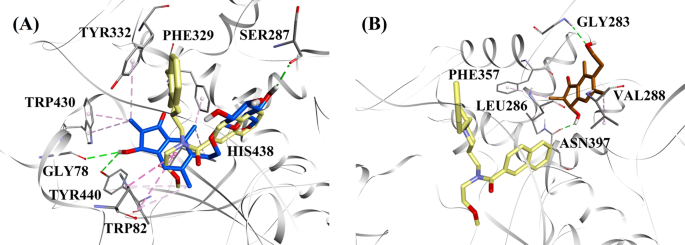
Moleculaire dockingmodellen voor a de gemengde-type BChE-remmer (2R,3R)-pteroside C (blauwe kleur) en b de niet-concurrerende BChE-remmer (2S,3R)-pterosine C (bruine kleur). De gedockte poses zijn gesuperponeerd op de X-straal kristalstructuur van 3F9 (gele kleur) (PDB code: 4TPK). BChE, actieve site residuen en verbindingen worden getoond door respectievelijk lint, lijn en stick modellen. De kleuren van de stippellijnen verklaren het type van de verschillende interacties: waterstofbruginteracties (groen) en hydrofobe interacties (roze). BChE butyrylcholinesterase
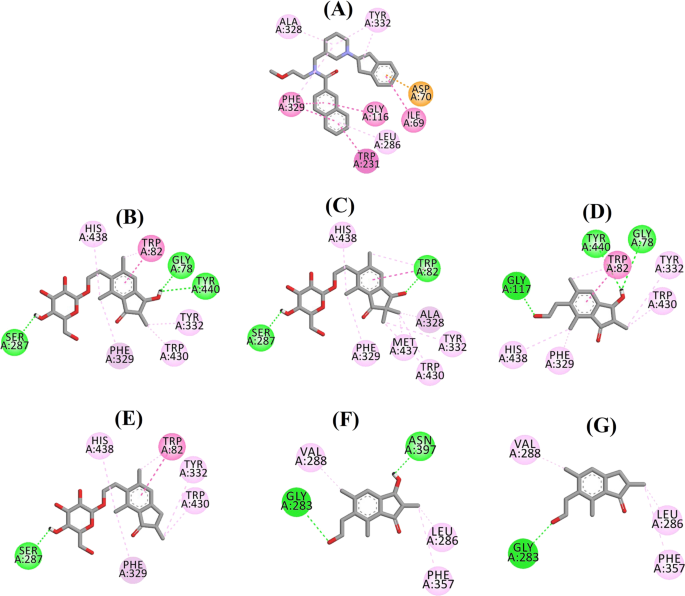
Ligandinteractiediagram van a 3F9, b (2R,3R)-pteroside C, c (3S)-pteroside D, d (2R,3R)-pterosine C, e (2R)-pteroside B, f (2S,3R)-pterosine C en g (2R)-pterosine B in de actieve site van BChE. De kleuren van de stippellijnen verklaren het type van de verschillende interacties: waterstofbruginteracties (groen), hydrofobe interacties (roze) en π-anion interacties (goud). BChE butyrylcholinesterase
De gedockte pose van 3F9 vertoonde een B.E. van -8,49 kcal/mol. Zoals weergegeven in Fig. 8a, waren hydrofobe interacties voornamelijk verantwoordelijk voor de ligandbinding. Ile69, Gly116, Trp231, Leu286, Ala328, Phe329, en Tyr332 waren verantwoordelijk voor de hydrofobische interacties, terwijl Asp70 een π-anion interactie vertoonde. Figuur 8b-e illustreert de gedokte poses van respectievelijk (2R,3R)-pteroside C, (3S)-pteroside D, (2R,3R)-pterosine C en (2R)-pteroside B (remmers van het gemengde type BChE). Hun bindingspocket was samengesteld uit Gly78, Trp82, Gly117, Ser287, Ala328, Phe329, Tyr332, Trp430, Met437, His438, en Tyr440. In overeenstemming met hun activiteitenniveaus vertoonden (2R,3R)-pteroside C (IC50 = 5,29 µM), (3S)-pteroside D (IC50 = 19,3 µM), (2R,3R)-pterosine C (IC50 = 20,3 µM) en (2R)-pteroside B (IC50 = 62,0 µM) een B.E. van respectievelijk -7,23, -6,59, -6,52, en -4,38 kcal/mol. Zoals aangetoond in Fig. 8b, vertoonde de 3-OH groep van de indanon ring van (2R,3R)-pteroside C twee waterstofbruggen met Gly78 en Tyr440 op afstanden van respectievelijk 2.87 en 2.89 Å. In het geval van (3S)-pteroside D beïnvloedde de 2,2-dimethylgroep aan de indanonring de bindingsinteracties van de 3-OH groep (Fig. 8c). De 3-OH groep vormde slechts één waterstofbrug met Trp82 op een afstand van 2,94 Å, wat resulteerde in een relatief lagere activiteit van (3S)-pteroside D dan (2R,3R)-pteroside C. Zoals te zien is in Fig. 8d, slaagde (2R,3R)-pterosine C er niet in een waterstofbrug te vormen met Ser287 door de afwezigheid van een 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolgroep. Bijgevolg leverde het een lagere activiteit op dan (2R,3R)-pteroside C. Het ontbreken van de OH-groep op positie 3 van de indanonring bleek verantwoordelijk te zijn voor de zeer lage activiteit van (2R)-pteroside B (Fig. 8e), dat er niet in slaagde waterstofbruggen te vormen met Gly78 en Tyr440 zoals (2R,3R)-pteroside C.
De gedockte poses van (2S,3R)-pterosine C en (2R)-pterosine B (niet-competitieve BChE remmers) worden respectievelijk getoond in Fig. 8f, g. Deze in het dock geplaatste poses werden ingesloten door Gly283, Leu286, Val288, Phe357, en Asn397. Wat hun activiteitsniveaus betreft, vertoonden (2S,3R)-pterosine C (IC50 = 44,3 µM) en (2R)-pterosine B (IC50 = 48,1 µM) een B.E. van respectievelijk -5,40 en -5,06 kcal/mol. (2S,3 R)-Pterosine C was krachtiger dan (2R)-pterosine B door de aanwezigheid van een extra OH groep op positie-3 van de indanon ring, die een waterstofbrug vormde met de CO groep van Asn397 op een afstand van 2.05 Å (Fig. 8f). Andere interacties bleken vergelijkbaar te zijn met (2R)-pterosine B. Zoals te zien in Fig. 8g, vormde de OH groep in de hydroxyethyl groep op positie-6 van de indanon ring een waterstofbrug met Gly283 op een afstand van 2.40 Å. De residuen Leu286, Val288, en Phe357 namen deel aan hydrofobe interacties.
Remmers van het gemengde type binden zowel aan het vrije enzym als aan het enzym-substraat complex, wat aangeeft dat deze verbindingen zich kunnen binden aan de katalytische plaats van elk corresponderend enzym. Niet-competitieve remmers binden zich aan de allosterische plaats van het vrije enzym of het enzym-substraatcomplex. Een recente studie suggereerde dat competitieve, mixed-type en niet-competitieve remmers verschillende plaatsen in de bindingszakken van BACE1, AChE, en BChE38 bezetten. Tijdens de docking voor de evaluatie van het remmende mechanisme van pterosinederivaten, werden de bindingsplaatsen van de verbindingen gedefinieerd volgens hun type van remming. De docking resultaten gaven aan dat de bindingsplaatsen van mixed-type en niet-competitieve remmers voor BACE1, AChE, en BChE elkaar gedeeltelijk overlappen op elke corresponderende actieve site en consistent waren met een eerder rapport38.
BBB permeabiliteit
PAMPA-BBB, een in vitro artificiële membraanpermeabiliteitsbepaling voor de BBB, is een van de meest betrouwbare fysisch-chemische screeninginstrumenten in de vroege fase van de ontdekking van geneesmiddelen gericht op het CZS40. Het PAMPA-BBB-systeem modelleert de transcellulaire passieve diffusie van chemicaliën over de BBB en meet de strikt passieve transportmechanismen via een kunstmatig lipidemembraan op de effectieve permeabiliteit (Pe, cm/s). Op basis van het patroon dat is vastgesteld voor de voorspelling van de BBB-permeatie, werden verbindingen ingedeeld in (i) “CNS+” (hoge BBB-permeatie voorspeld); Pe (10-6 cm/s) > 4,00, (ii) “CNS-” (lage BBB-permeatie voorspeld); Pe (10-6 cm/s) < 2,00, en (iii) “CNS+/-” (BBB-permeatie onzeker); Pe (10-6 cm/s) van 4,00 tot 2,00. Dienovereenkomstig vertoonden (2R)-pterosine B, (2S)-pterosine P, en (2S)-pterosine A een hoge BBB-permeatie met Pe-waarden van respectievelijk 60,3 × 10-6 cm/s, 7,92 × 10-6 cm/s, en 6,26 × 10-6 cm/s (tabel 4, aanvullende informatie 4). De Pe-waarde van (2R)-pterosine B was 1,7-maal hoger dan die van het CNS-medicijn verapamil (Pe = 34,6 × 10-6 cm/s), dat als positieve controle werd gebruikt. (2S,3R)-Pterosine C en (2R,3R)-pterosine C vertoonden een onzekere BBB-permeatie met Pe-waarden van respectievelijk 2,34 en 1,98. (2R,3R)-Pteroside C, (3S)-pteroside D, en (2R)-pteroside B, die van alle geteste pterosinederivaten de krachtigste BACE1- en cholinesteraseremmende activiteiten vertoonden, vertoonden een zeer lage BBB-permeabiliteit. De aanwezigheid van de 2-hydroxymethyl-tetrahydro-pyran-3,4,5-triolgroep zoals in pterosidederivaten resulteerde in een opmerkelijke afname van de BBB-permeabiliteit. Vergeleken met (2R)-pterosine B verminderde de aanvullende aanwezigheid van de OH-groep op positie-3 van de indanonring zoals in pterosine C, de hydroxymethylgroep op positie-2 van de indanonring zoals in (2S)-pterosine A of de hydroxymethylgroep op positie-5 van de indanonring zoals in (2S)-pterosine P ook de BBB-permeabiliteit aanzienlijk. Gezien de uitzonderlijk hoge BBB-permeabiliteit en de significante remming van BACE1, AChE en BChE, heeft (2R)-pterosine B wellicht het potentieel om een sterke anti-AD-activiteit te vertonen.
Effecten van (2R)-pterosine B en (2R,3R)-pteroside C op de secretie van Aβ-peptiden door neuronale cellen
Om de functie van (2R)-pterosine B en (2R,3R)-pteroside C in het verminderen van de uitscheiding van Aβ uit neuronale cellen, gebruikten wij een murine neuroblastoma cellijn die stably overexpresses menselijke APPswe. De cellijn is een cellulair model van AD gekenmerkt door de overmatige uitscheiding van Aβ40 en Aβ42. Giftige amyloïde oligomeren worden gevormd uit de twee isovormen van Aβ peptide met verschillende lengte. Aβ40 is de meest voorkomende Aβ-isovorm in de hersenen41, terwijl Aβ42 aanzienlijk toeneemt bij bepaalde vormen van AD42. Sandwich ELISA van Aβ40 toonde aan dat (2R)-pterosine B de hoeveelheid Aβ40 peptide die uit de neuroblastoma cellen in media werd uitgescheiden tot 50% verminderde bij 500 μM (P < 0,01) (Fig. 9a). Evenzo verminderde de secretie van Aβ42 peptide door de neuroblastoomcellen aanzienlijk in aanwezigheid van 500 µM van (2R,3R)-pteroside C (P < 0,05) (Fig. 9b). Concluderend kan worden gesteld dat (2R)-pterosine B en (2R,3R)-pteroside C de afscheiding van Aβ-peptiden uit neuroblastoomcellen aanzienlijk verminderden bij een concentratie van 500 μM.

a Effect van (2R)-pterosine B op de secretie van Aβ-peptiden. APPswe-secreterende neuroblastoomcellen werden gedurende 24 uur behandeld met 12, 60, 250 of 500 µM (2R)-pterosine B, waarna geconditioneerde media werden verzameld in aanwezigheid van een proteaseremmer. β-SI (10 µM) werd gebruikt als positieve controle. De negatieve controle omvatte cellen die gekweekt waren in afwezigheid van teststoffen. Kwantitatieve analyse van uitgescheiden Aβ40 en Aβ42 in de geconditioneerde media werd uitgevoerd met sandwich-ELISA. De gemiddelden ± SD van drie onafhankelijke experimenten worden getoond. De uitgescheiden Aβ40-peptiden namen significant af in aanwezigheid van 500 µM (2R)-pterosine B. CON negatieve controle, β-SI β-secretase inhibitor III, Aβ β-amyloïde. *P < 0.05, **P < 0.01. b Effect van (2R,3R)-pteroside C op de afscheiding van Aβ-peptiden. APPswe-secreterende neuroblastoomcellen werden behandeld met 12, 60, 125, 250, of 500 µM van (2R,3R)-pteroside C gedurende 24 uur, en geconditioneerde media werden verzameld in aanwezigheid van proteaseremmer. β-SI (10 µM) werd gebruikt als positieve controle. De negatieve controle omvatte cellen die gekweekt waren in afwezigheid van teststoffen. Kwantitatieve analyse van uitgescheiden Aβ40 en Aβ42 in de geconditioneerde media werd uitgevoerd met sandwich ELISA. De gemiddelden ± SD van drie onafhankelijke experimenten worden getoond. Het uitgescheiden Aβ42 peptide daalde significant in aanwezigheid van 500 µM (2R,3R)-pteroside C. CON negatieve controle, β-SI β-secretase inhibitor III, Aβ β-amyloïd. *P < 0.05
Cytotoxiciteit van pterosinederivaten op basis van MTT-test43
In het algemeen vertoonden pterosinederivaten een verwaarloosbare cytotoxiciteit tegen diverse normale en kankercellijnen, zoals SH-SY5Y humaan neuroblastoom, C6 rattengliacellen, NIH3T3 muizenembryofibroblasten en B16F10 muizenmelanoom, met LD50-waarden boven 0.5 mM (aanvullende informatie 5). In het bijzonder, (2R)-pterosine B, (3R)-pterosine D, (2S)-pterosine P, (2S)-pteroside A, (2R)-pteroside B, en (2R,3R)-pteroside C toonden geen cytotoxiciteit tegen de geteste cellijnen met LD50 waarden boven 5 mM. Intrigerend is dat verschillende pterosinen relatieve antiproliferatieve effecten vertoonden tegen SH-SY5Y neuronale cellen in vergelijking met C6 gliale cellen van mesenchymale oorsprong. De huidige resultaten waren consistent met een eerder rapport dat suggereerde dat pterosinederivaten niet toxisch zijn voor mensen26.