

De bijdrage van een zoutbrug aan de algehele stabiliteit van de gevouwen toestand van een eiwit kan worden beoordeeld met behulp van thermodynamische gegevens die zijn verkregen uit mutagenesestudies en kernspinresonantietechnieken. Met behulp van een gemuteerd pseudo-wild-type eiwit dat specifiek gemuteerd is om neerslag bij hoge pH te voorkomen, kan de bijdrage van de zoutbrug aan de totale vrije energie van de gevouwen eiwittoestand worden bepaald door een puntmutatie uit te voeren, waarbij de zoutbrug wordt veranderd en bijgevolg gebroken. Zo is in het T4 lysozym een zoutbrug gevonden tussen asparaginezuur (Asp) op residu 70 en een histidine (His) op residu 31 (figuur 3). Plaatsgerichte mutagenese met asparagine (Asn) (figuur 4) werd uitgevoerd, waardoor drie nieuwe mutanten werden verkregen: Asp70Asn His31 (mutant 1), Asp70 His31Asn (mutant 2), en Asp70Asn His31Asn (dubbelmutant).

Wanneer de mutanten zijn vastgesteld, kunnen twee methoden worden gebruikt om de vrije energie te berekenen die met een zoutbrug gepaard gaat. Een methode omvat de observatie van de smelttemperatuur van het wild-type eiwit versus die van de drie mutanten. De denaturatie kan worden gevolgd door een verandering in het circulaire dichroïsme. Een verlaging van de smelttemperatuur wijst op een vermindering van de stabiliteit. Dit wordt gekwantificeerd aan de hand van een door Becktel en Schellman beschreven methode, waarbij het verschil in vrije energie tussen de twee wordt berekend via ΔTΔS. Deze berekening levert enkele problemen op en kan alleen worden gebruikt met zeer nauwkeurige gegevens. In het voorbeeld van T4 lysozym was ΔS van het pseudo-wild-type eerder gerapporteerd bij pH 5,5, zodat het gemiddelde temperatuurverschil van 11 °C bij deze pH vermenigvuldigd met de gerapporteerde ΔS van 360 cal/(mol-K) (1,5 kJ/(mol-K)) een vrije-energieverandering oplevert van ongeveer -4 kcal/mol (-17 kJ/mol). Deze waarde komt overeen met de hoeveelheid vrije energie die door de zoutbrug wordt bijgedragen aan de stabiliteit van het eiwit.
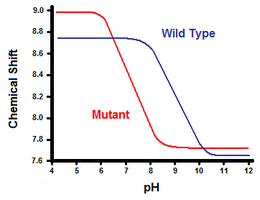
De tweede methode maakt gebruik van kernspinresonantiespectroscopie om de vrije energie van de zoutbrug te berekenen. Er wordt een titratie uitgevoerd, waarbij de chemische verschuiving wordt geregistreerd die overeenkomt met de protonen van de koolstof die aan de carboxylaat- of ammoniumgroep grenst. Het middelpunt van de titratiecurve komt overeen met de pKa, of de pH waarbij de verhouding geprotoneerde:gedeprotoneerde moleculen 1:1 is. Uitgaande van het voorbeeld van T4-lysozym wordt een titratiecurve verkregen door waarneming van een verschuiving van het C2-proton van histidine 31 (figuur 5). Figuur 5 toont de verschuiving in de titratiecurve tussen het wild-type en de mutant waarin Asp70 Asn is. De gevormde zoutbrug bevindt zich tussen het gedeprotoneerde Asp70 en het geprotoneerde His31. Deze interactie veroorzaakt de verschuiving in de pKa van His31. In het ongevouwen wildtype eiwit, waar de zoutbrug afwezig is, zou His31 een pKa van 6,8 hebben in H20 buffers van matige ionensterkte. Figuur 5 toont een pKa van het wild-type van 9,05. Dit verschil in pKa wordt ondersteund door de interactie van His31 met Asp70. Om de zoutbrug in stand te houden, zal His31 proberen zijn proton zo lang mogelijk vast te houden. Wanneer de zoutbrug wordt verbroken, zoals bij de mutant D70N, verschuift de pKa terug naar een waarde van 6,9, veel dichter bij die van His31 in ongevouwen toestand.
Het verschil in pKa kan worden gekwantificeerd om de bijdrage van de zoutbrug aan de vrije energie weer te geven. Met behulp van de vrije energie van Gibbs: ΔG = -RT ln(Keq), waarbij R de universele gasconstante is, T de temperatuur in Kelvin, en Keq de evenwichtsconstante van een reactie in evenwicht. De deprotonatie van His31 is een zure evenwichtsreactie met een speciale Keq die bekend staat als de zure dissociatieconstante, Ka: His31-H+ ⇌ His31 + H+. De pKa is dan als volgt aan Ka gerelateerd: pKa = -log(Ka). Het vrije-energieverschil tussen de mutant en het wild-type kan nu worden berekend met behulp van de vrije-energievergelijking, de definitie van pKa, de waargenomen pKa-waarden, en de relatie tussen natuurlijke logaritmen en logaritmen. In het voorbeeld van T4 lysozym leverde deze aanpak een berekende bijdrage van ongeveer 3 kcal/mol aan de totale vrije energie op. Een soortgelijke aanpak kan worden genomen met de andere deelnemer in de zoutbrug, zoals Asp70 in de T4 lysozym voorbeeld, door het controleren van de verschuiving in pKa na mutatie van His31.
Een woord van voorzichtigheid bij het kiezen van de juiste experiment betreft de locatie van de zoutbrug binnen het eiwit. Het milieu speelt een grote rol in de interactie. Bij hoge ionische sterktes, kan de zoutbrug volledig worden gemaskeerd omdat een elektrostatische interactie is betrokken. De His31-Asp70 zoutbrug in T4 lysozym was begraven binnen het eiwit. Entropie speelt een grotere rol bij zoutbruggen aan het oppervlak, waar residuen die normaal kunnen bewegen, worden ingesnoerd door hun elektrostatische interactie en waterstofbruggen. Het is aangetoond dat dit de entropie voldoende vermindert om de bijdrage van de interactie bijna uit te wissen. Oppervlaktezoutbruggen kunnen op soortgelijke wijze worden bestudeerd als begraven zoutbruggen, waarbij gebruik wordt gemaakt van dubbele mutantcycli en NMR-titraties. Hoewel er gevallen bestaan waarin begraven zoutbruggen bijdragen tot de stabiliteit, zoals alles, bestaan er uitzonderingen en kunnen begraven zoutbruggen een destabiliserend effect vertonen. Ook oppervlaktezoutbruggen kunnen, onder bepaalde omstandigheden, een stabiliserend effect hebben. Het stabiliserende of destabiliserende effect moet van geval tot geval worden beoordeeld en er kunnen weinig algemene uitspraken worden gedaan.